
-
Publish Your Research/Review Articles in our High Quality Journal for just USD $99*+Taxes( *T&C Apply)
Offer Ends On
Publish Your Research/Review Articles in our High Quality Journal for just USD $99*+Taxes( *T&C Apply)
Offer Ends On
Henry E. Young* and Oscar Tellez
Corresponding Author: Henry E. Young PhD, Chief Science Officer, Dragonfly Foundation for Research and Development, 12443 Venice Blvd (Corporate Office), Foley, AL 36535 USA.
Received: June 19, 2026 ; Revised: June 21, 2026 ; Accepted: June 22, 2026 ; Available Online: June 23, 2026
Citation: Young HE & Tellez O. (2026) Introduction to Adult Telomerase Positive Stem Cells (aTPSCs). J Stem Cell Ther Res, 1(1): 1-52.
Copyrights: ©2026 Young HE & Tellez O. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Views & Citations
Likes & Shares
The field of regenerative medicine has long sought the “Holy Grail”, a cell that has unlimited proliferation potential, can differentiate into any cell, can restore dead and dying cells to normal functional cells, and can be used for anyone, making them a universal regenerative stem cell. Many types of stem cells have been suggested to be the Holy Grail. Most notable are three categories of stem cells that have been widely studied since 1990. 1. Embryonic stem cells (ESCs) that are isolated from the inner cell mass of developing embryos, 2. Mesenchymal stem cells (MSCs) that were originally isolated from bone marrow of post-natal adults, and 3. Induced pluripotent stem cells (iPSCs) that are derived by transfecting embryonic genes, e.g., Oct-4, SOX2, c-Myc, and Klf4, into an adult differentiated cell, most notably adult dermal fibroblasts. Each one has advantages and disadvantages. Both ESCs and iPSCS, because of the presence of the telomerase enzyme have unlimited proliferation potential. Both ESCs and iPSCs are very plastic, they can differentiate into any somatic cell of the body. Unless prevented to do so though, both ESCs and iPSCs will form teratomas (cancerous cells) by spontaneous differentiation. To prevent teratoma formation, both ESCs and iPSCs need to be pre-differentiated, which in the process, loses their plasticity for forming multiple cells. The ESCs, by virtue of being isolated from the inner cell mass of developing embryos, are allogeneic, expressing self-recognition molecules which will induce a graft versus host disease (GvHD) response in the recipient. The iPSCs, being isolated from the same individual, were thought to negate the GvHD response. Unfortunately, the transfection process alters the self-recognition molecules to an extent to make them initiate a GvHD response. The MSCs have a limited lifespan of 70 population doublings before they senesce and die. MSCs, like all telomerase negative progenitor cells, decrease in number with increasing age of the individual. MSCs only form fat, cartilage, and bone, and therefore are not plastic in the ability to form all somatic cell types. Autologous (same person) MSCs do not elicit a GvHD, whereas (donor) allogeneic MSCs induce a GvHD due to presence of MHC Class-1 self-recognition cell surface molecules. In contrast to the above, we would like to offer a fourth category of stem cells for consideration, endogenous adult telomerase positive stem cells (aTPSCs). The aTPSCs retain the telomerase enzyme after birth that endows them unlimited proliferation potential. They are present throughout the lifespan of the individual. Collectively, they will form any cell of the conceptus, including all somatic cells of the body, gender-specific gametes, the nucleus pulposus of the intervertebral disc, and extraembryonic membranes, placenta and umbilical cord. Their default state is that of a dormant, quiescent, hibernating cell. They have to the stimulated by biological agents to do anything, hence, no teratoma formation because there is no spontaneous differentiation. They are very tightly controlled with respect to function: proliferation, progression, induction, and anti-differentiation.
Keywords: Adult, Telomerase Positive, Totipotent, Pluripotent, MSCs, ESCs, iPSCs
PODCAST - 1
If adult salamanders can regenerate tissues, why can’t humans?
Can you take us back to that moment?
Why did you ask that question, and why did it become important for your career?
Actually, the initial question for my Master’s thesis was: if juvenile salamanders can regenerate a limb, why can’t adult salamanders?
I read the existing literature on the subject at that time, circa 1950-1973 [1-12]. The multiple groups that had studied the phenomenon of limb regeneration in aquatic juvenile and adult newts and aquatic salamanders kept them at 4oC water, and fed beef liver daily during daylight hours. They assayed limb regeneration every five days for 30 days (newts) to 45 days (salamanders) for 6-9 time points. The investigators concluded that the juvenile aquatic salamanders damaged tissues dedifferentiated into the blastema and regenerated the limb [2].
They also concluded that limb regeneration did not occur in adult salamanders, which they kept under the same conditions and viewed them at the same time points.
From their observations they concluded that adult salamanders had lost the ability to regenerate a limb.
So, for my study, I kept the adult salamanders under the same environmental conditions as the juvenile aquatic species: 4oC water and fed beef liver daily during daylight hours. Using these environmental conditions, all my adult salamanders died of starvation before I could even start my experiments. So, I went to my Chairman and asked how I should proceed.
He said “what do you know of your model system?” – answer “they are adult salamanders”.With a twinkle in his eyes, he said “you need to dig deeper and find out everything you can about your model system”From my own field work observations and further literature research, I discovered that adult salamanders:
1. Are terrestrial, not aquatic, they hide in burrows in the ground during daylight hours.
2. They come out at night looking for a nocturnal (active at night) food source, preferably one that moves on its own.
3. Their preferred food source is cockroaches, although they were also preferential to night crawlers.
4. Migrate during the first cold rain of the fall, they spawn to the ponds they were born in to breed (copulate) with the opposite gender.
5. They did not eat when they were breeding.
6. When breeding was concluded, they return to their burrows
When I presented my findings to my Chairman, he told me four things that have followed me throughout my research career.
1. Know you model system
2. Tissue NEVER lies
3. You need to understand what your model system is telling you, and act accordingly
4. Just because something hasn’t been reported, does not mean it doesn’t exist, all it means is that it hasn’t been discovered yet.
So, I built a large, deep, terraria for all the salamanders, similar to environmental conditions in the wild. I released night crawlers into their terraria every two days. They apparently liked their conditions because weights increased. I repeated the experiment with fat, happy, and sassy salamanders. I kept the same observation times: every five days. But because it took the adult terrestrial salamanders over 370+ days to fully regenerate a limb, I was making 74+ observations, depending on the particular species in the genus Ambystoma (maculatum, annulatum, tigranum, and texanum).
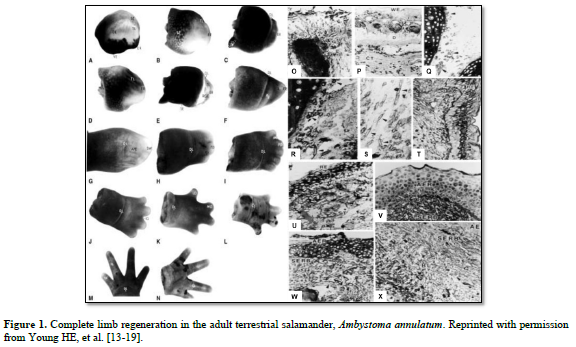
And I discovered some interesting points (Figure 1):
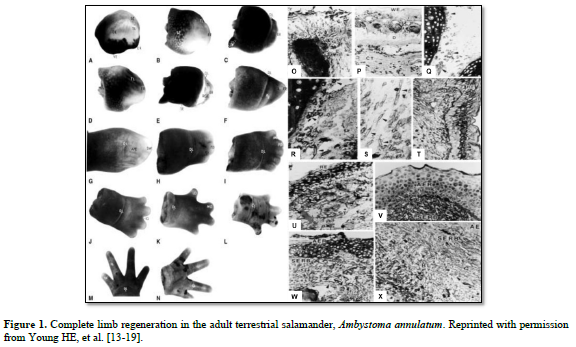
There was the same series of events that occurred in each of the four adult salamanders I examined, Ambystoma maculatum, annulatum, texanum, tigranum:
1. After amputation of the limb a transitional scar formed covering the wound site, basically a band-aid separating a very hostile external environment from a very delicate internal environment (O). Next, there was the appearance of macrophages that appeared underneath the transitional scar and cleaned out all the debris and dying cells, making the wound area sterile (P). There was formation of an apical epidermal ridge of non-descript cells on top of the transitional scar (B, C).
2. This ridge of cells began secreting a concentration gradient of sulfated, carboxylated, and neutral glycoproteins into the area through and underneath the transitional scar (U & V).
3. Previously very small unobserved cells, now covered in halos of heparan sulfate-PGs (HS-PG) (Q), broke loose from the more proximal connective tissues of the dermis; periosteum; perichondrium; muscle endomysium, perimysium, epimysium; nerve endoneurium, perineurium, and epineurium; and connective tissues surrounding the vasculature (R,S) and migrated to an area beneath the AER (T), eventually forming the sub-epidermal ridge blastema (SERB) (T- X)
4. The very small cells shed the HS-PG coverings and formed an indistinct mass of very small cells (T).
5. Then the very small cells they began to proliferate and physically push the AER outward (B-L).
6. This formed a gradient of differentiated tissues: differentiated tissues of the non-transected limb, through intermediaries similar to same tissues during embryonic development, to non-distinct cells of the “blastema”.
7. As the length of the appendage increased, the more proximal intermediaries turned into adult differentiated tissues.
Bottom line from those experiments:
Adult terrestrial salamanders regenerate perfectly fine, if given the appropriate environment, diet, exercise (hunting for food), and sleep cycle.
So, from knowledge of what occurred in adult salamanders spawned the question:
If adult salamanders can regenerate tissues, why can’t humans?
Or better yet, are we keeping humans under the wrong environmental conditions for regeneration to occur?
Why did you ask that question, and why did it become important for your career?
My parents and a close family member, had/have serious genetically inherited and acquired health issues. These health issues included heart disease and diabetes (father), Hashimoto’s disease, Systemic Lupus Erythematosus, Pulmonary Fibrosis, SLE-induced glaucoma, and SLE-induced Dementia (mother), and all of the above ‘inherited’ from both parents (close family member). Plus, he was diagnosed with Autoimmune Constellation Syndrome. To have this diagnosis one has to have a minimum of five autoimmune or autoimmune associated diseases. He has over 30. To give you an idea of what he had to deal with since he was 4 years old (in order of appearance): Hashimoto’s disease maintaining a short stature (4’5” tall) and overweight to severely obese (250-350 lbs.) from 4-17 years of age; Sjogren’s disease; Scleroderma; Alopecia; multiple allergies to foods, apparel, smoke, environment; adult respiratory distress syndrome (ARDS), pericarditis; pleuritis; pulmonary fibrosis; Rhinitis; Esophagitis; Tracheitis; Gastritis; Ileitis; Celiac Disease; Colitis; Rectifies (severe inflammation within the rectum); Hepatitis; Rhabdomyositis (severe inflammation of skeletal muscle), Rhabdomyolysis (wasting of skeletal muscle, think directed sarcopenia); Pancreatitis; Cholecystitis (Gall Bladder); Nephritis; Vasculitis; Systemic Lupus Erythematosus; osteopenia/osteoporosis (long term prednisone use), torsion (spiral) fracture of left leg into multiple pieces (due to a twisting fall); sterility; extreme sensitivity to sun light (photosensitivity) with resulting formation of keratoses; Neuropathies; Bi-Lateral Sciatica; Migraines; Cluster Headaches (Suicide Headaches); fibrosed CNs L1-S5 to his vertebral column (he said it felt like he was growing a dorsal fin from his vertebral column outward); extreme unrelenting pain (with the following pain killers given simultaneously every four hours: 64-mg of hydromorphone, 4x max dose Gabapentin, 2x max dose Baclofen, maxed out 12-hr Tylenol, Aspirin, Ibuprofen, and Naproxen, did not give him any relief from the pain); OIC (Opioid-induced constipation); TIAs (transient ischemic attacks, mini strokes); Cardiomyopathies; Tachycardia inducing Heart attacks, Autoimmune-induced Type-1 Diabetes; Chronic Kidney Disease; Rheumatoid Arthritis; Atrial Fibrillation; SLE-induced glaucoma; and cataracts. Suffice it to say, in the 70 years that he has been expressing various autoimmune and associated diseases, he states that it has been an interesting journey [20].
If these previously unrecognized primitive cells that I discovered in adult salamanders were also present in humans, could I restore the health of individuals in my family? That started my 50+ year quest.
2. Why are Salamanders so important?
Salamanders are the highest order of animal that will completely regenerate a limb that is an exact duplicate of the histoarchitecture of the limb that was lost.
What can they do that makes scientists ask bigger questions about healing and regeneration?
Scientists can ask about genetic control of regeneration; where are the genes that control the process; is epigenetics involved; is methylation involved; how are the biological clocks of various organs related to regeneration; are components of the ECM (extracellular matrix, e.g., collagens, proteoglycans, glycoproteins) involved in the process, and if so, how; will the cells involved spontaneously form a limb or is it a tightly controlled; are there biological factors involved that control the process; where are those factors located; how do they interface with the primitive cells; what are the characteristics of these cells; what techniques can you use to identify them; so on and so forth; and lastly, are similar scenarios and components present in humans. If so, how can they be used to restore damaged tissues in humans.
For example, for my PhD degree I performed glycoconjugate histochemistry on serial sections of the regenerating limb tissues to identify particular proteoglycans (PGs): chondroitin sulfate-PG, keratan sulfate-PG, dermatan sulfate-PG, chondroitin sulfate/keratan sulfate-PG (also called Aggrecan), non-sulfate chondroitin-PG, hyaluronic acid, sulfated glycoproteins, neutral glycoproteins, and carboxylated glycoproteins, and using microspectrophotometry, quantify their amounts. I also used scanning electron microscopy coupled with glycoconjugate histochemistry and X-ray energy dispersive microanalysis to quantity ECM components. What I discovered was that each tissue in the body, be it fully differentiated, newly forming, partially regenerated, or regenerated, had a unique glycoconjugate profile, which I called its “fingerprint”. From there, I wanted to be able to isolate these glycoconjugates in a biologically active form to see what effects they would have on the aTPSCs. After obtaining my PhD degree (1984), I obtained a postdoctoral fellowship in a laboratory that performed glycoconjugate biochemistry to isolate and characterize proteoglycans within the ECM [21-23].
3. Human Regeneration Question
What does regeneration mean to you?
Regeneration to me means restoration of the damaged and/or missing cells and tissues recreating the normal histoarchitecture of the lost tissues, and thereby restoring normal function.
4. Early Scientific Environment
When you began exploring this idea, what was the scientific field focused on?
Were people open to the idea that adult humans might still have still powerful cells, or was that idea outside the mainstream?
Basically, it was outside mainstream thought processes. This was because true adult stem cells had not yet been discovered. Therefore, common belief, based of dogma, said that these particular stem cells did not exist.
To be able to receive government funding to study any phenomenon, one of my PhD mentors told us that the game plan for receiving NIH funding was to do the experiments ahead of time, but wait to publish. Write the experiments performed as an application for a grant; including hypothesis, M&Ms, costs, etc. and submit. When that grant was funded, work on experiments for the next grant submission. At termination period of the first grant, publish the results. So, you would have fulfilled what you set out to prove, or disprove. In addition, it was far better to have a “story” to tell using multiple technologies, then to perfect a single technology and using the same technology on multiple tissues. So, my “story” for my research career has been the role of aTPSCs in regenerative medicine.
I isolated aTPSCs from chickens, cloned them from single cells using conditioned medium, characterized them, etc., and submitted the grant to NIH (circa 1989).
My “pink sheet” response from the grant reviewers was “well written grant, but your data is flawed. Everyone knows (dogma) that adult stem cells don’t exist. … But if they did exist, you would need to show them in a research animal which is preferably a mammal, not a chicken. We would suggest a mouse”.
I published the chicken methodology data: ELICA and Isolation protocols to a third-tier journal, Journal of Tissue Culture Methods. With respect to this particular journal, one would submit their manuscript to the editor of the journal. The editor would send your manuscript to one of the reviewers to repeat your experiments exactly as written. If they could not repeat your experiments and get exactly the same results, the manuscript was either rejected outright or revised significantly to match the permutations of your methodologies to get the experiments to work. Both manuscripts were accepted without revision [24,25].
I then started using mice: Balb-C (standard research mouse) and CBF-1 (NIH’s aging model) as my research subjects and repeated the experiments. The lifespan of a Balb-C mouse is 24 months (equivalent to about 60 years of age), whereas the lifespan of a CBF-1 mouse is 36-40 months (equivalent to about 120 years of age, the pre-programmed limit for humans) [21-23].
I joined Dr. Arnold Caplan’s lab in 1984 (nine years after I had discovered the aTPSCs in adult salamanders) for a postdoctoral fellowship in glycoconjugate biochemistry. Again, I wanted to isolate the glycoconjugates in their biological active form so I could apply them to the aTPSCs to determine if they were involved in cellular regeneration.
Being a biochemist by training, Dr. Caplan was a “lumper” with respect to anatomical structures. Being an anatomist/histologist/histochemist by training, I was a “splitter” with respect to anatomical structures. So, while Dr. Caplan viewed skeletal muscle as a single organ, I viewed skeletal muscle as a collection of individual tissues. There are three levels of structural elements composing skeletal muscle the organ. The first level is composed of mature myotubes having myosatellite cells (myoblast progenitor cells) outside their plasma membrane, but inside their basement membrane (consisting of type-IV collagen, entactin, nidogen, insoluble fibronectin, etc.), each myotube was surrounded by loose fibrous connective tissue (type-1 and type-12 collagens. Type-12 collagen is the bridge molecules between type-1 collagen and its associated GPs and PGs of the ECM) termed the endomysium. Embedded within the endomysium were capillaries, aTPSCs, hyaluronic acid with attached CS-PGs. Collections of these myotube structures were bundled together to form fascicles, the second level. The connective tissue surrounding the bundled fascicles were a moderately dense fibrous connective tissue (type-1 and -12 collagens) called the perimysium. Embedded within the perimysium are arterioles, venioles, small lymphatic vessels, motor end plates, sensory muscle spindles, nerve fibers, aTPSCs, and hyaluronic acid with attached CS-PGs. At the third level, bundles of myotubes came together to form skeletal muscle the organ, surrounded by a dense regular connective tissue covering termed the epimysium (type-1 and -12 collagens). Contained within the epimysium were muscular arteries, muscular veins, lymphatics, nerve fibers, Golgi tendon organs, aTPSCs, and hyaluronic acid with attached CS-PGs. The epimysium is continuous with tendons (connecting adjacent muscles to each other or connecting muscle to bones) [21-23].
I learned to isolate and characterize extracellular matrix PGs from Dr. David Carrino in Dr. Caplan’s lab [23] and glycoproteins from Dr. Masaki Yanagashita during a visit to Dr. Vince Hascall’s lab at NIH NIDR.
I submitted my next NIH grant (1990) dealing with aTPSCs in mice, Balb-C (normal) and CBF-1 (aging). It was denied, because Dogma says that adult stem cells do not exist. But if adult stem cells did exist, the reviewers stated that I would need to show them in a larger mammal, such as a rat.
I submitted manuscripts concerning aTPSCs in the chicken [24,25] and in both normal age mice and aged mouse models [21-23]to tier one journals: Cell, Nature, Science, PNAS (Proceedings of the National Academy of Science, USA). They were either rejected outright because of Dogma – adult stem cells do not exist; or stuck in review for over two years.
I had named the telomerase positive MesoSCs in that original paper “adult mesenchymal stem cells”, because they could form 37 separate and unique cell types within the embryonic mesodermal lineage (mesenchyme).
The manuscripts were finally released back to me after Dr. Arnold Caplan published his seminal “adult mesenchymal stem cell” paper (Caplan AI. Mesenchymal stem cells. J Orthop Res. 1991; 9:641-650 [26]) showing the discovery of an adult stem cell isolated from bone marrow that would form 3 cell types: fat, cartilage, and bone. After his publication, my submitted manuscripts for chicken and mouse endogenous stem cells [24,25] were finally returned rejected for not being novel with respect to adult mesenchymal stem cells.
I finally published our work with chicken and mouse telomerase positive mesodermal stem cells (TP-MesoSCs), calling them “adult pluripotent mesenchymal stem cells” because of their ability to form 37 separate cell types [27-30]. I should have called them mesodermal stem cells, but I wanted to demonstrate a distinct difference between Caplan’s mesenchymal stem cells which would only three cell types (white fat, hyaline cartilage, and intramembranous bone) [31,32] and my telomerase positive MesoSCs, which would form 37 distinct cell types to several tier-2 journals [26-28]. Again, in some articles I originally called them pluripotent mesenchymal stem cells for their ability to form 37 different cell types within the mesodermal lineage, instead of just three cell types formed by Caplan’s (telomerase negative) tripotent progenitor MSCs, fat, cartilage, and bone [26,31,32].
Based on Caplan’s seminal adult MSC paper [26], adult stem cells were finally acknowledged to exist. But they had short comings. They had a defined lifespan of 70 population doublings before they senesced and died; they decreased in number with increasing age of the individual; and they would only form three cell types, e.g., (white) fat, (hyaline) cartilage, and (intramembranous) bone [26,30-32]. Around the same time as Caplan’s MSC publication [26], other “adult stem cell” papers were published: adult neural stem cells [33-39], adult hematopoietic stem cells [40-46], adult liver stem cells [47-52], adult pancreatic stem cells [53 -57], adult lung cells [58-63], etc. The organ specific adult stem cells correlated with Caplan’s MSCs, e.g., their lifespan conformed to Hayflick’s Limit of 70 population doublings from birth, they decreased with increasing age of the individual, and they only formed organ-specific cell types.
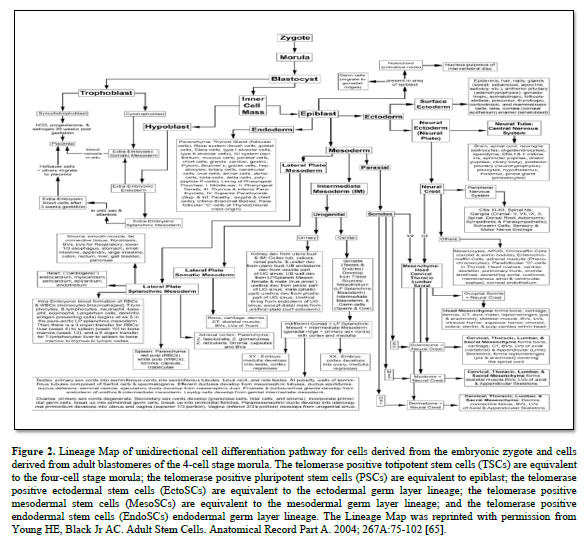
In 1998, Dr Thomson published on the derivation human embryonic stem cells from human blastocysts (ESCs). These ESCs were isolated from the inner cell mass of developing embryo and were pluripotent in that they could form any somatic cell of the body [64]. They were equivalent to the differentiation potential of the inner cell mass of the developing embryo (Figure. 2) [65].
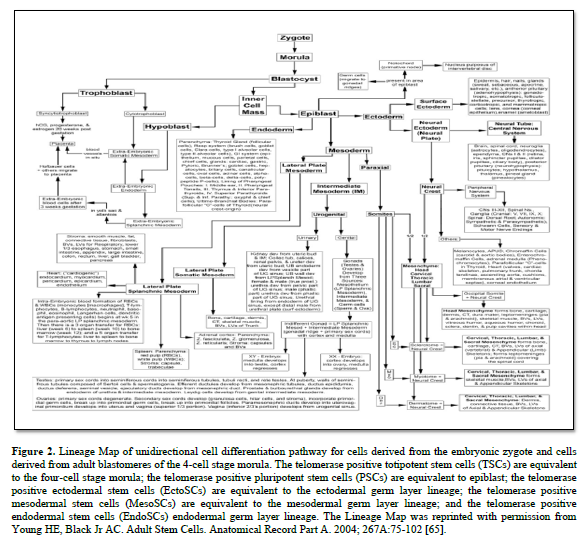
ESCs also contained the telomerase enzyme, which allows them unlimited proliferation potential [66-69]. Unfortunately, there were significant problems with ESCs when growing them outside the body (ex vivo) or transplanting them as naïve cells into the body. Unless they were physically prevented from differentiating with leukemia inhibitory factor (LIF) [70-77]; or stimulate to premature differentiation [78-82], ESCs will spontaneously differentiate into all somatic cells of an individual [83,84]. If the ESCs are in the uterus, they complete their normal pre-programming differentiation process forming an individual [84-86]. If they are outside the uterus, the ESCs will form a mass of somatic cells with no defined structure, called a teratoma cancer [87-91].
So now the politics. On one hand there were cell specific adult stem cells, that conformed to Hayflick’s Limit of 70 population doublings before they would senesce and die. They deceased with increasing age of the individual. Their differentiation was growth factor driven, but they weren’t as plastic as naïve ESCs [26,31,32,33-63].
On the other hand, ESCs would form every somatic cell of the body, proving to be very plastic. Containing the telomerase enzyme ESCs had essentially an unlimited proliferation potential. But, being allogeneic, they would express self-recognition markers of the donor and stimulate a GvHD, as well as forming teratomas if implanted in their naïve state [64-91].
So political debate was centered on ESCs versus MSCs, which one was better.
All the while we were publishing with collaborators in 2nd and 3rd tier journals on the aTPSCs [92-131]. As well as starting preclinical animal models of diseases: Parkinson’s disease [114], myocardial infarction [106], pulmonary fibrosis [107], and self-renewing immunoprotected pancreatic islet organoids for type-1 diabetes [110].
The political fall-out about using human embryos to derive embryonic stem cells lasted until Yamanaka (2009) published his seminal work on induced pluripotent stem cells (iPSCs) [132-134]. He placed embryonic genes (Oct-4, Sox2, c-Myc, and Klf4) into adult differentiated cells to mimic ESCs. And he did it so well with the transfection that the iPSCs expressed the same attributes as ESCs. The iPSCs expressed the telomerase enzyme [135-145], having an unlimited proliferation potential. In the naïve state, their inherent plasticity was their ability to form any somatic cell type in the body, which occurred spontaneously, just like ESCs [146-150]. This spontaneous differentiation occurred anywhere, in the culture dish, in an organism, etc., forming a teratoma (cancerous tissue) [146-150]. Unfortunately, to keep teratomas from forming they needed to pre-differentiate the cells into a single cell type [146-148]. By pre-differentiating the iPSCs or ESCs, they lose the naïve plasticity that made them a stem cell of choice for the Holy Grail.
5. What kept you curious?
You have spent decades studying this field. What kept you committed to this kind of research when most of the stem cell field was focused in other directions?
My early work with chickens, mice, and rats, demonstrated a very unique population of cells, with all the positives of ESCs and iPSCs, and “adult stem (progenitor cells)”, but none of the negatives:
1.Telomerase positive, so unlimited proliferationpotential as long as they stay uncommitted to aparticular lineage
2.Present throughout the lifespan of the individual
3.Found within connective tissue niches throughoutthe body
4.Will form literally any cell type in the body, e.g.,all somatic cells, gender-specific gametes, nucleuspulposus of intervertebral disc, and extraembryonicmembranes, placenta and umbilical cord
5.Proliferation is biological agent driven
6.Differentiation is biological agent driven
7.Anti-differentiation is biological agent driven
8.Once committed, progression is biological agentdriven
We have shown this same activity in 15 species of animals, including humans: amphibians (four species of adult terrestrial salamanders), reptiles (Komodo Dragon), avians (chickens and Wadel Crane), mice (Balb-c, CBF-1), rats (outbred Sprague-Dawley, inbred Wistar-Furth), rabbits, cats, dogs, sheep, goats, pigs, cows, bear (spectacled), horses, and humans (newborn to late geriatric) [151].
As I stated previously, this area is very personal to me. My family members had/have serious acquired and genetic health issues. If these previously unrecognized cells were present in humans, could I restore the health of my parents and myself, and in the process everyone else?
To achieve that goal, I like to think backwards (reverse chronological order) from my end goal, that gives me a straight-line pathway from start to finish:
End Goal: treating humans (and animals) with gender-matched universal aTPSCs world-wide.
24.Wide-spread treatment
23.FDA approval for commercialization
22.Testing CNSP vs Fresh isolate aTPSCs vs TSCs Ex vivo–determine safety & efficacy
21.Apply for IND from FDA for clinical trials
20.Clinical trials of Ex vivo propagated TP-TSCS – provesafety and efficacy
19.Apply for IND from FDA for clinical trials
18.Propagation of universal TP-TSCs Ex vivo
17.Clinical trials: Fresh isolate aTPSCs & CNSP to proveefficacy
16.Apply for IND from FDA for clinical trials
15.Treatment in humans (my family members).
14.Wide-spread IRB-approved clinical trials to prove safety(and efficacy)
13.Focused IRB-approved clinical trials to prove safety andefficacy
12.Pre-clinical animal models of disease
11.Characterization studies
10.Biobanking, Storage and Cryopreservation
9.Effects of biological agents on clones of aTPSCs
8.Generation of cell-specific exosomes,
7.Genomic labeling to track cells in vitro and in situ
6.Repetitive Single cell clonogenic analysis
5.Cell sorting
4.Cell surface marker profiles
3.Propagation
2.Plating
1.Isolation
Since FDA allows experimentation on oneself without reprimand, I was the first to receive an autologous transplant of aTPSCs (systemic delivery). My HIPPA code number is HM00001. My mind set at the time was, if the technology failed, I would be dead and the technology would not move forward.
My group started IRB approved compassionate use clinical trials in 2010. First in Parkinson patients, and then in COPD, IPF, and cardiomyopathy patients, matching the pre-clinical animal model systems [152-156].
I was also the first to receive a gender-matched, ABO blood group-matched allogeneic aTPSCs, by directed delivery and IV delivery. My mind set was the same, if the technology failed, I would be dead, and it would not move forward.
Positive results from the first trials allowed us to expand into other diseases: terminal, chronic diseases with no known cures, traumatic injuries, chronic orthopedic problems, autoimmune diseases, neurodegenerative, pulmonary, cardiovascular, and systemic [157-172].
It was too late to treat my father, because he passed away from a heart attack while I was still characterizing the cells. And it was too late to treat my mother, because she passed away from pulmonary fibrosis and dementia secondary to SLE while we were doing the preclinical animal studies. But I was just in time to treat my other family member. We had just started the IRB-approved compassionate use clinical trials for Parkinson’s disease and pulmonary diseases (COPD and IPF). I remember my PCP (board certified family physician) coming to me during a break in the phase tutorials, we both taught in the same phase. During break he put his arm around me and said “Henry, I know what you do for your research, go do it on yourself.” “Why?” “You have barely two weeks to live. You have already lost two organ systems and the remaining systems are operating at less than 25%. Your body is shutting down. You will be dead within two weeks if not sooner if you don’t save yourself.”
That night I discussed the situation with my wife. The next day I went to my chairman with letter in hand “As you know I have some serious health issues, I need about two weeks to go and get treated. If you don’t agree, here is my letter of resignation”. He said “Go for it and your job will be waiting for you when you return”.
And my wife did the same with her employer. “You know Henry is sick. He needs treatment or he will die. I need time off to take him to get treated. If you don’t agree, here is my letter of resignation”. They agreed as well.
In April of 2011 I had my first full autologous aTPSC transplant. Right after that first transplant, I was euphoric, absolutely no pain anywhere, I felt like Superman. The next day I woke up depressed, the extreme unrelenting pain was back. “Someone give me a gun I want to shoot myself”. The second day after treatment I woke up “Hey, this is strange, less pain than yesterday”. Third day same, less pain than day before. By the 7th day after treatment, I was neuropathically pain free, and basically have been ever since. But after a month, while there was no further downward progression of organ failure, I didn’t get any better. So, I had the first of nine allogeneic gender-matched, ABO-blood group-matched (3)and O-negative (6) aTPSC transplants. Those have beeninterspersed with 20 total autologous aTPSC transplants.With the allogeneic transplants, my signs and symptoms ofneurodegenerative diseases, cardiovascular morbidities,pulmonary fibrosis, chronic kidney disease, celiac disease,and SLE-associated morbidities began to reverse and myorgan functions began to increase. I topped out at levels thatwere 70% normal for a 20-year-old (acceptable to me).
My current mind set is, if my technologies, using either autologous and/or allogeneic aTPSCs, can bring my family member back from my death bed and give him a reasonable quality of life, then the aTPSCs should help people with other health problems as well.
Part 2. The Stem Cell Categories Most People Know
6.The Three Main Categories
At a high level can you explain what those three categories are:
ESCs – embryonic stem cells are derived from inner cell mass of embryo, they are pluripotent in their ability to form all somatic cell types of the body, they contain the telomerase enzyme for essentially unlimited proliferation potential. Initially published for humans in 1998 by Dr James Thomson [64].
iPSCs – induced pluripotent stem cells were generated by taking differentiated adult cells and transfecting into their nucleus four embryonic genes (Oct-4, SOX2, c-Myc, and Klf4) to have them mimic embryonic stem cells: pluripotent in ability to form all somatic cell types of the body, contains telomerase enzyme for essentially unlimited proliferation potential. Published by Yamanaka in 2009 [132].
MSCs – an “adult stem cell” (actually a tripotent progenitor cell) originally derived from bone marrow that will form three differentiated cell types: fat, cartilage, and bone. MSCs are telomerase negative. They have a lifespan of 70 population doublings before they senesce and die. MSCs decrease with increasing age of the individual. Published by Arnold Caplan in 1991 [26].
7.Embryonic Stem Cells
What makes embryonic stem cells so important clinically?
Embryonic stem cells were originally designed to study embryogenesis in utero: discovering genes and teratogens impacting signaling pathways, differentiation steps, etc., to determine how one could repair, for example, inborn errors of metabolism, spina bifida, Chiari syndrome, microcephaly, autism, cleft lip, cleft palate, etc., etc., etc., before the baby was born.
Then someone had the “bright idea” that they could use ESCs in adults (post-natal individuals) to repair acquired and genetic diseases.
What limitations have made them difficult to use clinically?
1.First, and foremost, is Politics – “killing an embryoto acquire ESCs. Embryos have rights too”.
2.Obtaining funding from the government because ofthe above to study ESCs.
3.Their spontaneous differentiation into multiple celltypes, necessitates using an inhibitory agent (e.g.,LIF) to prevent spontaneous differentiation.
4.ESCs formation of teratomas when transplanted invivo in a naïve state.
5.ESCs needed to be pre-differentiated beforetransplant to prevent teratoma formation.
6.ESCs are allogeneic (non-self).
7.ESCs express self-recognition cell surfacemolecules that will induce a graft versus hostdisease response in the recipient, HLA-DR markersfor hematopoietic lineage markers and MHC Class-1 markers for somatic cells that were not in thehematopoietic lineage.
8.Induced Pluripotent Stem cells
Can you explain what iPSCs are in simple terms?
The induced pluripotent stem cells were generated by taking differentiated adult cells, originally dermal fibroblasts/fibrocytes, but other cell types have been used as well. And using adenoviruses, transfecting four embryonic genes (Oct-4, SOX2, c-Myc, and Klf4) into the nucleus of the adult differentiated cells, to have them mimic embryonic stem cells. After which, they were pluripotent in ability to form all somatic cell types of the body, and expressed the telomerase enzyme for essentially unlimited proliferation potential.
Why was that discovery such a big deal?
iPSCs gave scientists a method to reprogrammed cells to a less differentiated cell type, e.g., a pluripotent cell that would form all somatic cells of the body. Since they were from the person’s own body the self-recognition cell surface markers would be the same so there would be no graft versus host disease (GvHD) response (theory).
9.Limits of Reprogramming
What are some of the challenges the field still has to solve?
Are there issues around safety, consistency, tumor risk, or clinical practicality?
Several of the major problems of iPSCs is that they mimic ESCs too well and have demonstrated the same inherent problems:
1.They form teratomas when transplanted in vivoin a naïve state.
2.Since they spontaneously differentiate intomultiple cell types, this necessitates using aninhibitory agent (LIF or some facsimile) to prevent spontaneous differentiation.
3.They need to be pre-differentiated to preventtumor formation.
4.And even though they come from the sameindividual, the reprogramming changes theexpression of the self-recognition molecules on their cell surfaces making them seem allogeneic to the recipient’s immune system, which will induce a graft versus host disease response in the recipient, destroying the iPSCs.
5.Labs are propagating the iPSCs at a doubling ratefaster than their cell cycle rate to increasenumber of cells generated. Unfortunately, as the doubling rate increases the number of mutations formed increases, and begins to increase exponentially at 10^9 cells.
6.Permanently mutated cells can have deleteriouseffects downstream in the treatment phase.
7.Only correct non-mutated iPSCs need/should tobe selected for human treatments.
8.It takes about 6-12 months to isolate, propagate,induce, select, and generate sufficient numbersof specific iPSC cell types for transplant. Increasing costs with respect to time, reagents, etc.
9.And lastly, from my own observations andresearch: the “body” does not likedifferentiated cell types, it views them as foreign, even those expressing the same MHC Class-1 markers. The body will wall them off from the rest of itself and encapsulates it with scar tissue. It prefers an undifferentiated cell that it can manipulate and dictate what it becomes.
10.Mesenchymal Stem Cells
MSCs have become the most talked about cell types in regenerative medicine.
What are MSCs?
And what do you think they can and cannot do?
Since previous to Caplan’s publication in the journal Science, adult stem cells were thought NOT to exist (Dogma). Now, here is a paper from a known scientist (biochemist) that says that ADULT STEM CELLS do exist in the form of mesenchymal stem cells. And that these adult stem cells can be isolated from adult bone marrow, and will form fat, cartilage, and bone.
Through my early years of my research, when I characterized aTPSCs (TSCs, PSCs, EctoSCs, MesoSCs, and EndoSCs), I also characterized the tripotent MSC, Caplan’s MSC. I characterized mixed isolates of aTPSCs and MSCs, clones of all six cell types derived from single cells derived by repetitive single cell clonogenic analyses, and from genomically-labeled aTPSCs clones compared to the unlabeled clone of MSCs. In this last instance, I sent the clones of aTPSCs and MSCs to Cecille Duplaa at INSERM in France to genomically label the cells. She tried to transfect all the clones with the Lac-Z gene for beta-galactosidase, but instead of using adenoviruses [scientifically accepted method, but sometimes the viruses can go “rogue” especially in long term culture of cells], she used lipofectin. Lipofectin “transfects” the cells during cell division. The more the cells divide during a given time frame the higher percentage of cells are transfected. The transfection rate for TSCs were 99%; for PSCs 98%; for EctoSCs, MesoSCs, and EndoSCs, greater than 95%; and for MS <5%.
With Arnold Caplan being the “lumper” that he was, and me being the “splitter” that I am as well as being a trained histologist/glycoconjugate histochemist/immunocytochemist, I also characterized the cell fates of his tripotent progenitor MSC, e.g., “fat (white fat), cartilage (hyaline cartilage), bone (intramembranous bone)”, using morphological, histochemical, and immunocytochemical criteria [130,131,151,173-175].
Table 1. Antibodies, Immunocytochemistry, & Histochemistry for Phenotypic Expression Markers
| Antibody | Antigen | Embryological Origin |
| CEA-CAM-1 | Carcinoembryonic antigen-cell adhesion molecule-1 | Totipotent |
| HCEA | Human Carcinoembryonic antigen | Totipotent |
| CEA | Carcinoembryonic antigen | Totipotent |
| CD66e | Carcinoembryonic antigen | Totipotent |
| DH-TuAg1 | Spermatogonia | Totipotent Gamete |
| MC-480 | SSEA-1 | Pluripotent |
| MC-631 | SSEA-3 | Pluripotent |
| MC-813 | SSEA-4 | Pluripotent |
| CD10 | Neutral endopeptidase | Pluripotent |
| AlkPhos | Alkaline Phosphatase | Pluripotent |
| CD56 | Neural cell adhesion molecule | Ectoderm |
| Pax-6 | Neurogenic lineage | Ectoderm |
| FORSE-1 | Neuronal precursor cells | Ectoderm |
| Vimentin | Cells of neurogenic lineage | Ectoderm |
| Nestin | Cells of neurogenic lineage | Ectoderm |
| R401 | Nestin-neuronal lineage | Ectoderm |
| HNES | Nestin-neuronal lineage | Ectoderm |
| MAB353 | Nestin-neuronal lineage | Ectoderm |
| RT-97 | Neurofilaments = neurons | Ectoderm |
| NF68 | Neurofilament-68 = neurons | Ectoderm |
| S-100 | Neurofilaments-100 = neurons | Ectoderm |
| NF-145 | Neurofilaments-145 = neurons | Ectoderm |
| N-200 | Neurofilaments-200 = neurons | Ectoderm |
| 8A2 | Neurons | Ectoderm |
| NG2 | Neurons | Ectoderm |
| TH | Tyrosine hydroxylase, precursor to neural transmitters | Ectoderm |
| SV2 | Synaptic vesicles | Ectoderm |
| DOPA | Dopamine, transmitter of dopaminergic neurons | Ectoderm |
| T8660 | Beta-tubulin-III | Ectoderm |
| Tuj1 | Beta-tubulin | Ectoderm |
| GFAP | Glial-fibrillary acidic protein | Ectoderm |
| CNPase | Glial cells = oligodendrocytes & astrocytes | Ectoderm |
| Rip | Oligodendrocytes | Ectoderm |
| MOSP | Oligodendrocyte specific proteins | Ectoderm |
| MAB | Oligodendrocyte marker | Ectoderm |
| 40E-C | Radial cells and radial glial cells | Ectoderm |
| VM-1 | Keratinocytes | Ectoderm |
| M3F7 | Type-IV collagen, basement membrane | Ectoderm & Mesoderm |
| 31-2 | Laminin, basement membrane | Ectoderm & Mesoderm |
| 5D2-27 | Cell adhesion molecule | Ectoderm & Mesoderm |
| B3/D6 | Fibronectin, basement membrane | Ectoderm & Mesoderm |
| 5C6 | Type-IV collagen, basemen membrane | Ectoderm & Mesoderm |
| Anti-type IV | Type-IV collagen | Ectoderm & Mesoderm |
| 33-2 | Heparan sulfate proteoglycan | Ectoderm & Mesoderm |
| Anti-HSPG | Heparan Sulfate proteoglycan | Ectoderm & Mesoderm |
| 5D4 | Keratan sulfate proteoglycan | Ectoderm & Mesoderm |
| 2E8 | Laminin, basement membrane | Ectoderm & Mesoderm |
| D3 | Desmin, in all 3 muscle groups | Ectoderm & Mesoderm |
| Anti-vimentin | Vimentin, lens of the eye | Ectoderm & Mesoderm |
| D76 | Desmin, in all 3 muscle groups | Ectoderm & Mesoderm |
| CD13 | Amino endopeptidase | Mesoderm |
| 12/101 | Skeletal Muscle | Mesoderm |
| C3/1 | Glycoprotein of myoblast plasma membrane | Mesoderm |
| OP-137 | MyoD | Mesoderm |
| F5D | Myogenin = skeletal muscle | Mesoderm |
| ALD-66 | Slow twitch muscle fibers | Mesoderm |
| MF-1 | Fast twitch muscle fibers | Mesoderm |
| MF-5 | Myosin light chain-2 of fast muscle | Mesoderm |
| MF-20 | Sarcomeric myosin = skeletal muscle | Mesoderm |
| MF-30 | Neonatal and adult myosin | Mesoderm |
| ALD58 | Myosin heavy chain | Mesoderm |
| CH1 | Myosin tropomyosin | Mesoderm |
| A4.74 | Myosin fast chain | Mesoderm |
| JLA-20 | Actin | Mesoderm |
| Anti-Myosin | Skeletal muscle myosin | Mesoderm |
| IA4 | Smooth muscle alpha actin = smooth muscle | Mesoderm |
| Calp | Calponin | Mesoderm |
| MAB-3252 | Cardiotin = cardiac myocytes | Mesoderm |
| MAB1548 | Myosin heavy chain of cardiac muscle | Mesoderm |
| M-38 | Type 1 collagen | Mesoderm |
| SP1.D8 | Procollagen type-III | Mesoderm |
| Anti-type-II | Type-II collagen | Mesoderm |
| WV1D1 | Bone sialoprotein II = bone | Mesoderm |
| Anti-OsteC | Osteocalcin / Bone Gla-protein | Mesoderm |
| MP111 | Osteopontine = bone | Mesoderm |
| Von Kossa | Stain calcium in bone | Mesoderm |
| EGTA | Leaches Calcium from bone, negative control | Mesoderm |
| CIIC1 | Type-II collagen | Mesoderm |
| II-4CII | Type-II collagen | Mesoderm |
| Anti-type2 | Type-II collagen | Mesoderm |
| HC-II | Human type-II collagen | Mesoderm |
| D1-9 | Type-IX collagen = cartilage | Mesoderm |
| 9/30 | Cartilage link protein | Mesoderm |
| 12/21 | Cartilage proteoglycan hyaluronate binding region | Mesoderm |
| 12C5 | Versican hyaluronate binding region | Mesoderm |
| H-DC34 | Sialomucin-containing hematopoietic/endothelial cells | Mesoderm |
| CD31 | PECAM, Peripheral endothelial cell adhesion molecule | Mesoderm |
| P1H12 | Human endothelial cell surface marker | Mesoderm |
| P2B1 | Peripheral endothelial adhesion molecule | Mesoderm |
| P8B1 | VCAM, vascular cell adhesion molecule | Mesoderm |
| P2H3 | CD62e, E-selectin (vasculature) | Mesoderm |
| H-endo | CD146, Endothelial cells | Mesoderm |
| H5A4 | CD11b, granulocytes, monocytes, NK-cells | Mesoderm |
| H4C4 | CD44, hyaluronate receptor | Mesoderm |
| Hermes-1 | CD44, hyaluronate receptor | Mesoderm |
| H5A5 | CD45, all hematopoietic cells except RBCs | Mesoderm |
| H5C6 | CD63, macrophages, monocytes, platelets | Mesoderm |
| HFSP | Human fibroblast specific protein | Mesoderm |
| 1B10 | Fibroblast-specific protein | Mesoderm |
| Sudan Black-B | Stains fat (adipocytes) | Mesoderm |
| Oil Red-O | Stains fat (adipocytes) | Mesoderm |
| H-AFP | Human alpha-fetoprotein = fetal liver | Endoderm |
| R-AFP | Rat alpha-fetoprotein = fetal liver | Endoderm |
| DESMO | Endodermal epithelial marker of liver | Endoderm |
| LAP | Canalicular cell surface protein of liver | Endoderm |
| 151-Ig | Liver epithelial growth factor | Endoderm |
| HA4c19 | Bile canalicular cells of liver | Endoderm |
| OC2 | Progenitor cells, oval cell, & biliary cells of liver | Endoderm |
| OC3 | Progenitor cells & biliary cells of liver | Endoderm |
| OC4 | Progenitor cells & biliary cells of liver | Endoderm |
| OC5 | Progenitor cells & biliary cells of liver | Endoderm |
| OC10 | Progenitor cells & biliary cells of liver | Endoderm |
| H.4 | Intracellular staining of liver hepatocytes | Endoderm |
| H.1 | Liver hepatocytes cell surface marker | Endoderm |
| DPPIV | Progenitor, canalicular, and biliary cells of the liver | Endoderm |
| OV6 | Biliary and oval cells of liver; biliary cells of liver | Endoderm |
| HESA | Human GI (Gastrointestinal) Epithelium | Endoderm |
| YM-PS087 | Glucagon-secreting cells of endocrine pancreas | Endoderm |
| YM-PS5088 | Insulin-secreting cells of endocrine pancreas | Endoderm |
| 11180 | Somatostatin-secreting cells of the endocrine pancreas | Endoderm |
| CK-19 | Ductal cells of the exocrine pancreas | Endoderm |
| ABL-93 | Lysosomal membrane glycoprotein | Ectoderm, Mesoderm, Endoderm |
| 22/18 | Regeneration cells | Ectoderm, Mesoderm, Endoderm |
| Telom | Telomerase positive cells | aTPSCs |
| CD90 | Glycosylphosphatidylinositol anchoring membrane protein (Thy-1) | Transition: EctoSCs, MesoSCs, and EndoSCs to Progenitor Cells |
| Thy-1 | Glycosylphosphatidylinositol anchoring membrane protein (CD90) | Transition: EctoSCs, MesoSCs, and EndoSCs to Progenitor Cells(e.g., MSCs) |
| CD95 | Cells undergoing apoptosis | Dead Cells |
| PI | Propidium Iodide, measure of live cells, flow cytometry | Live Cells |
| DAPI | Fluorescent marker to visualize living and fixed DNA | Live & Dead Cells |
| Gal-19 | Insect beta-galactosidase genomic marker | Cell tracking marker |
| Mallory Heidenhain One-Step | Identifies various cell types by color:Type-1 collagen – dark blueType-2 collagen – light blueSkeletal muscle – dark magentaCardiac muscle – intermediate magentaSmooth muscle – light magentaAdipose Tissue - whiteNerve fibers – lavenderRBCs - golden | Cells & Extracellular Matrix |
| Alcian Blue | Stains anions on carbohydrate & sulfate groups | Extracellular Matrix |
| AB 1.0 | Alcian Blue, pH 1.0 stains sulfate groups on GAGs | Extracellular Matrix |
| AB 2.5 | Alcian Blue, pH 2.5 stains carboxyl groups on GAGs | Extracellular Matrix |
| Alcec Blue | Stains anions on carbohydrate groups | Extracellular Matrix |
| AcB 1.0 | Alcec Blue, pH 1.0 stains sulfate groups on GAGs | Extracellular Matrix |
| AcB 2.5 | Alcec Blue, pH 2.5 stains carboxyl groups on GAGs | Extracellular Matrix |
| Safranin-O | Stains anions on carbohydrate & sulfate groups | Extracellular Matrix |
| SO 1.0 | Safranin-O, pH 1.0 stains sulfate groups on GAGs | Extracellular Matrix |
| SO 2.5 | Safranin-O, pH 2.5 stains carboxyl groups on GAGs | Extracellular Matrix |
| Enzyme | Streptomyces Hyaluronidase, negative staining control to verify presence of hyaluronic acid | Extracellular Matrix |
| Enzyme | Chondroitinase-AC, negative staining control to verify presence of chondroitin sulfate proteoglycans | Extracellular Matrix |
| Enzyme | Chondroitinase-ABC, negative control to verify presence of chondroitin non-sulfated proteoglycans | Extracellular Matrix |
| Enzyme | Keratanase, negative control to verify presence of keratan sulfate proteoglycans | Extracellular Matrix |
| Enzyme | Heparanase, to verify presence of heparan sulfate proteoglycans | Basement Membranes |
| PAS | Periodic Acid Schiff reaction for glycoproteins with vicinal hydroxyl groups | Extracellular Matrix |
Table 1. Immunocytochemistry with antibodies for cell-specific phenotypic expression markers and glycoconjugate histochemistry to determine “fingerprints” of specific cell types. Reprinted with permission from Young HE. A high throughput screening assay to quantify, visualize, and standardize biological activities: Enzyme-Linked Immuno-Culture Assay (ELICA). GSC Advanced Research and Reviews. 2025; 24(02): 091-114 [173]; Young HE, Speight MO. Osteoarthritis Treated with Telomerase-Positive Adult Stem Cells in Animals and Humans. Stem Cells Regen Med. 2020; 4(2):1-11 [158].
1. MSCs, specifically, form unilocular white fat. There are two types of fat: unilocular white fat and multilocular brown fat.
a. White fat
i. A single large vesicle is present filling the cytoplasm
ii. It has a single laterally-located nucleus
1. Lipid within the vesicle stains with oil-loving dyes, such as Oil Red-O and Sudan Black-B
b. Multilocular Brown
i. Is multilocular, having a centrally-located nucleus
ii. Multiple small vesicles are contained within the cytoplasm
iii. Lipid within the vesicle(s) stains with oil-loving dyes, such as Oil Red-O and Sudan Black-B.
2. MSCs, specifically, will form hyaline cartilage. There are five types of cartilage in the body:
a. Fibrocartilage
i. Appearance: Herringbone pattern of parallel dense regular connective tissue composed of collagen fibers, chondrocyte present within large oval-shaped lacunae
1. Located in symphysis pubis, menisci, labrum, and annulus fibrosis of intervertebral disc
ii. Collagen fibers are type-1 and type-12 collagen (bridge molecule)
1. Antibodies: M-38,
2. Mallory Heidenhain One Step – dark blue
3. Pure chondroitin sulfate proteoglycans
4. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
iii. Hyaluronic acid
1. Antibodies: H4C4 (CD44), Hermes-1 (CD44), 12C5 (HA binding region)
2. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Streptomyces hyaluronidase
b. Growth Plate Cartilage:
i. Location: metaphyseal portion of developing long bones during endochondral ossification and in hard callus during fracture repair
ii. Appearance: spicules composed of cartilage cores covered with lamellar bone, chondrocytes present in large oval lacunae, osteocytes present in small irregularly shaped lacunae
iii. Inner Cartilage cores
1. Collagens type-2 and type-9 (bridge molecule)
a. Antibodies: CIIC1, II-4CII, Anti-type2, Anti-type-II, HC-II,D1-9, 9/30, 12/21, 12C5
b. Mallory Heidenhain One Step: - light blue
2. Chondroitin sulfate/keratan sulfate proteoglycans (Aggrecan)
a. Glycoconjugate Histochemistry:
i. Chondroitin Sulfate Glycosaminoglycan chains
1. Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
ii. Keratan Sulfate Glycosaminoglycan chains
1. Alcian Blue pH 1.0, Alcec Blue pH 1.0, Safranin-O pH 1.0 with and without Keratanase
3. Hyaluronic acid
a. Antibodies: H4C4 (CD44), Hermes-1 (CD44), 12C5 (HA binding region)
b. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Streptomyces hyaluronidase
iv. Outer layers of Lamellar bone
1. Collagen fibers are type-1 and type-12 collagen (bridge molecule)
a. Antibodies: M-38,
b. Mallory Heidenhain One Step – dark blue
c. Pure chondroitin sulfate proteoglycans
d. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
2. Hyaluronic acid
a. Antibodies: H4C4 (CD44), Hermes-1 (CD44), 12C5 (HA binding region)
b. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Streptomyces hyaluronidase
c. Hyaline cartilage,
i. Location: attachment of ribs to sternum
ii. Appearance: random pattern of chondrocytes within large oval lacunae embedded with an amorphous extracellular matrix
iii. Extracellular Matrix
1. Collagens type-2 and type-9 (bridge molecule)
a. Antibodies: CIIC1, II-4CII, Anti-type2, Anti-type-II, HC-II,D1-9, 9/30, 12/21, 12C5
b. Mallory Heidenhain One Step: - light blue
2. Chondroitin sulfate/keratan sulfate proteoglycans (Aggrecan)
a. Glycoconjugate Histochemistry:
i. Chondroitin Sulfate Glycosaminoglycan chains
1. Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
ii. Keratan Sulfate Glycosaminoglycan chains
1. Alcian Blue pH 1.0, Alcec Blue pH 1.0, Safranin-O pH 1.0 with and without Keratanase
3. Hyaluronic acid
a. Antibodies: H4C4 (CD44), Hermes-1 (CD44), 12C5 (HA binding region)
b. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Streptomyces hyaluronidase
d. Elastic cartilage,
i. Location: Pinna of ear, eustachian tube, epiglottis
ii. Appearance: random pattern of chondrocytes within large oval lacunae embedded with an amorphous extracellular matrix containing elastic fibers
iii. Extracellular Matrix
1. Elastic fibers – elastin stain
2. Collagens type-2 and type-9 (bridge molecule)
a. Antibodies: CIIC1, II-4CII, Anti-type2, Anti-type-II, HC-II, D1-9, 9/30, 12/21, 12C5
b. Mallory Heidenhain One Step: - light blue
3. Chondroitin sulfate/keratan sulfate proteoglycans (Aggrecan)
a. Glycoconjugate Histochemistry:
i. Chondroitin Sulfate Glycosaminoglycan chains
1. Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
ii. Keratan Sulfate Glycosaminoglycan chains
1. Alcian Blue pH 1.0, Alcec Blue pH 1.0, Safranin-O pH 1.0 with and without Keratanase
4. Hyaluronic acid
a. Antibodies: H4C4 (CD44), Hermes-1 (CD44), 12C5 (HA binding region)
b. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Streptomyces hyaluronidase
e. Articular cartilage
i. Location: covering surface of articulating bones
ii. Appearance: 5 zones
1. Tangential zone
a. Type-2 and type-9 collagen fibers run parallel to surface,
b. Collagen fibers attach to lubricin,
i. A highly sulfated glycoprotein,
ii. forms a dipole with water,
iii. provides lubrication for articular joint
c. ECM consists of keratan sulfate proteoglycans only
2. Transitional zone
a. Type-2 and type-9 collagen fibers form crisscross pattern of fibers,
b. ECM consists of chondroitin sulfate proteoglycans only
3. Radial zone
a. Type-2 and type-9 collagen fibers run perpendicular to surface,
b. ECM consists of chondroitin sulfate/keratan sulfate proteoglycans attached to hyaluronic acid through a link protein (Aggrecan)
4. Tidewater Mark
a. Acellular
b. ECM consists of a jumbled mix of type-2 and type-9 collagen fibers, and a mix of chondroitin sulfate proteoglycans, keratan sulfate proteoglycans, and Aggrecan = chondroitin sulfate/keratan sulfate proteoglycans attached to hyaluronic acid through a link protein
5. Calcified cartilage
a. Chondrocytes present in intermediate-sized oval lacunae.
b. Type-2, type-9, type-1 and type-12 collagen fibers in a haphazard arrangement
c. ECM consists of Aggrecan = chondroitin sulfate/keratan sulfate proteoglycans attached to hyaluronic acid through a link protein, and calcium
d. Amorphous calcium phosphate
iii. Extracellular Matrix
1. Type-2 and type-9 (bridge molecule)
a. Antibodies: CIIC1, II-4CII, Anti-type2, Anti-type-II, HC-II, D1-9, 9/30, 12/21, 12C5
b. Mallory Heidenhain One Step: - light blue
2. Type-1 and type-12 collagen (bridge molecule)
a. Antibodies: M-38,
b. Mallory Heidenhain One Step – dark blue
3. Pure chondroitin sulfate proteoglycans
a. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
4. Pure keratan sulfate proteoglycans
a. Alcian Blue pH 1.0, Alcec Blue 1.0, Safranin-O pH 1.0 with and without Keratanase
5. Chondroitin sulfate/keratan sulfate proteoglycans (Aggrecan) double stained: first at pH 1.0 (Safranin-O) followed by pH 2.5 (Alcian blue or Alcec blue)
a. Chondroitin Sulfate Glycosaminoglycan chains
i. Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Chondroitinase-AC
b. Keratan Sulfate Glycosaminoglycan chains
i. Alcian Blue pH 1.0, Alcec Blue pH 1.0, Safranin-O pH 1.0 with and without Keratanase
c. Chondroitin sulfate/Keratan sulfate proteoglycan (Aggrecan)
i. Safranin-O at pH 1.0 followed by Alcian Blue (or Alcec Blue) at pH 2.5 with and without combined Chondroitinase-AC and Keratanase
d. Hyaluronic acid
i. Antibodies: H4C4 (CD44), Hermes-1 (CD44), 12C5 (HA binding region)
ii. Glycoconjugate Histochemistry: Alcian Blue pH 2.5, Alcec Blue 2.5, Safranin-O pH 2.5 with and without Streptomyces hyaluronidase
e. Amorphous calcium
i. Von Kossa stain with and without ethylene-glycine tetraacetic acid, a specific chelator for calcium
3. MSCs, specifically, will form intramembranous bone. During embryogenesis, bone forms by two mechanisms:
a. Endochondral ossification: mesodermal to cartilage model to bone, ex.; long bones, bones of face
i. Mesodermal cells
ii. Cartilage Model = growth plate cartilage (see above)
iii. Lamellar Bone
1. Heidenhain staining: Dark blue
2. Antibodies: M-38
3. Collagens: type-1 and type-12
4. Proteoglycans: Chondroitin sulfate-PG
b. Intramembranous ossification [direct mesoderm to bone]. ex., flat bones found in the calvaria, and flat portion of scapula.
i. Mesodermal cells
1. Lamellar Bone
2. Heidenhain staining: Dark blue
3. Antibodies: M-38
4. Collagens: type-1 and type-12
5. Proteoglycans: Chondroitin sulfate-PG
4. The first reported MSCs were a tripotent progenitor cell forming three cell types, white fat, hyaline cartilage, and intramembranous bone.
5. They are absent the telomerase enzyme.
6. They have a lifespan of 70 population doublings before they senesce and die.
7. They decrease in number with increasing age of the individual
8. The tripotent progenitor MSCs express CD90, CD105, CD123, CD166, and MHC Class-1 cell surface markers
9. Allogeneic MSCs when transplanted will induce a GvHD
a. If individual has competent immune system, it will kill the MSCs
b. If individual is immunocompromised, MSCs may kill the individual
10. Since tripotent progenitor mesenchymal stem cells did not live up to the promise of articular cartilage repair, Caplan changed the name of the tripotent MSCs to medicinal secretory cells (MSCs) [176-179].
a. These cells were proposed to release paracrine factors to modulate inflammation during the regenerative process
b. The medicinal MSCs express CD73, CD90, CD105, and MHC Class-1 cell surface markers [180-183].
c. Since the cell surface markers are not identical, medicinal MSCs are clearly a different cell type than the tripotent MSC
d. If used autologously, maybe function is correct
e. If used allogeneically, MAY induce GvHD, because of conflicting MHC Class-1 self-recognition molecules
Why MSCs became so popular
Why do you think MSCs became such a dominant topic in regenerative medicine?
Was it because they were easier to obtain, easier to study, safer, or more commercially practical?
1. Political
Initially, I believe it gave politicians something to compare to ESCs.
Table 2. Comparison of ESCs to MSCs
| Attributes | ESCs | MSCs |
| Plasticity | Any somatic cell | Fat, Cartilage, Bone |
| Telomerase enzyme | Present | Absent |
| Proliferation Potential | Unlimited | 70 Population Doublings |
| Age of Individual | Embryo | Adult |
| Location | Inner Cell Mass of Developing Embryo | Adipose Tissue, Bone Marrow, Wisdom Teeth, Umbilical Cord,Umbilical Cord Blood, any organ/tissue with a connective tissue compartment and associated with white fat, hyaline cartilage, and intramembranous bone |
| Self-Recognition Molecules | Allogeneic | Autologous,Allogeneic |
| Graft Vs Host Disease | Yes | No – AutologousYes – Allogeneic |
| Default State | Spontaneous Differentiation | Quiescent |
| Controlled by External Entities | No | Yes |
| Teratoma FormationIn Naïve State | Yes | No |
| Pre-Differentiate | Yes, otherwise Teratoma Formation | No |
| Easier to Study | No | Yes |
| Easier to Process Cells | No | Yes |
| Propagating Ex Vivo for transplants | Yes, but need LIF to prevent premature differentiation | Yes |
11. The Holy Grail ProblemWhy has the field struggled to find one perfect stem cell category?
Except for one group of cells, which we will get to in a few moments, no isolated cells to date filled all parameters for “Holy Grail” Wish List.
Table 3. Holy Grail Wish List
| Parameters | ESCs/iPSCs | MSCs |
| Telomerase Positive | Yes | No |
| Unlimited proliferation Potential | Yes | No |
| Present throughout lifespan of individual | NA | Decrease with increasing age |
| Absent Self-Recognition Molecules | No | No |
| Invisible to Immune System | No | No |
| Will form any somatic cell type | Yes | No,fat, cartilage, bone |
| Does NOT spontaneously differentiate | No | Yes |
| Pre-differentiation is NOT needed | No | Yes |
| Will NOT form teratomas | No | Yes |
| Function controlled by biological agents | No | Yes |
| Homing receptors for damaged cells | ?? | Yes |
| Naïve state forms what is lost or damaged | ?? | Yes |
| Does NOT overgrow existing cells/tissues | ?? | Yes |
| Exosome Production | Yes | Yes |
| Currently, can be propagated to large numbers without mutations | No | No |
| Universal stem cell transplant | No | No |
| Days of shelf-life at 4oC | ?? | ?? |
| Cryopreserved | -196 oC | -196 oC |
| Recovery Viability | ?? | 95% |
| Can be Freeze Dried | No | No |
| Restoration Viability | No | No |
| Can withstand-196 oC to +200 oC | NYD | NYD |
| Bio-printed into 3D Constructs for transplant | NYD | NYD |
Part 3: Introducing aTPSCs as a Fourth Category
1. What is an adult telomerase positive stem cell?
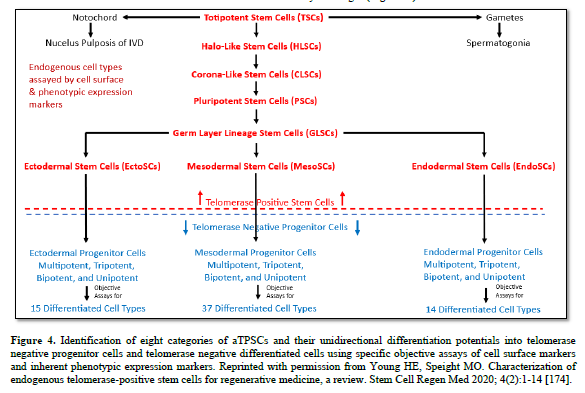
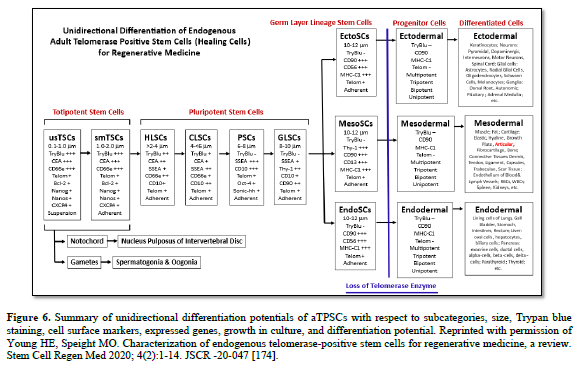
The aTPSCs are actually a category of cells (8 total, divided into 5 subcategories) that are found within all the connective tissues of the body after birth. They are unique in that they retain the telomerase enzyme after birth of the individual. This is unlike differentiated cells and progenitor cells that lose the telomeres enzyme at birth [184]. The aTPSCs are preprogrammed to heal/replace damaged tissues. Their default state in the body is as a dormant quiescent hibernating cell within the connective tissues, where they are maintained throughout the lifespan of the individual. The germ layer lineage stem cells: EctoSCs, MesoSCs, and EndoSCs, prefer an aerobic environment (21% Oxygen saturation) and are located very close to capillaries. TSCs and PSCs prefer an anaerobic environment (5% Oxygen saturation) and are located at maximum distance from the blood vessels.
The aTPSCs only become activated when they receive a signal (chemokine) released from damaged tissues. At which point they begin dividing symmetrically, forming daughter cells. The daughter cells undergo reverse diapedesis into the bloodstream. From there they home to the damaged tissues following an increasing concentration gradient of chemokines. Once on site, local metalloproteinases remove the blocking molecules from their receptor sites and they respond to locally released cues (exosomes/secretomes/biological agents) to restore whatever tissue is lost (Table 4).
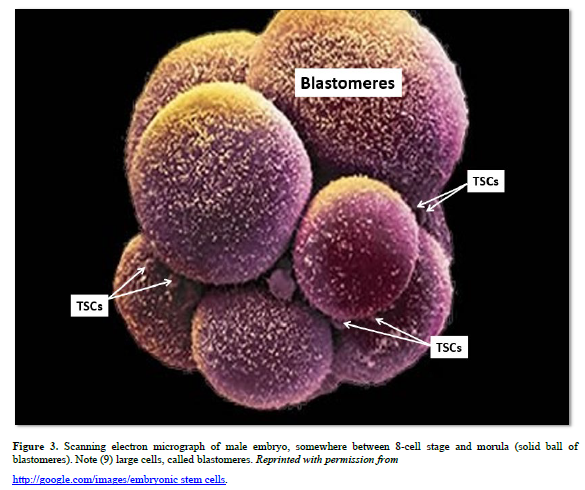
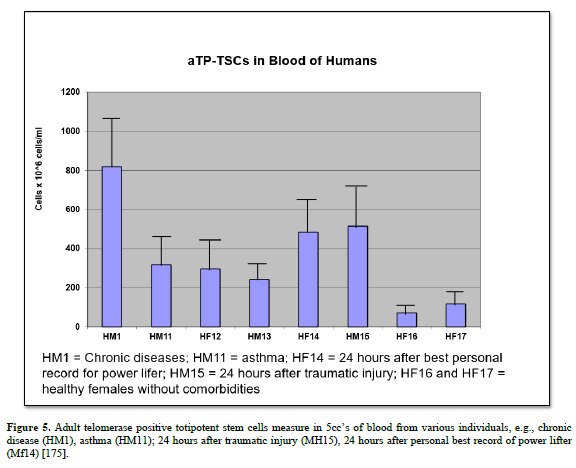

1. Farber J. An experimental analysis of regional organization in the regenerate forelimb of the axolotl (Ambystoma mexicanum). Arch Biol. 1959; 71:1-72.
2. Hay ED. Electron microscopic observations of muscle dedifferentiation in regenerating Amblystoma limbs. Dev Biol. 1959;1(6):555–585.
3. Gross J, Lapiere CM. Collagenolytic activity in amphibian tissues: a tissue culture assay. Proc Natl Acad Sci U S A. 1962 Jun;48(6):1014–1022.
4. Thornton CS. Amphibian limb regeneration. In: Advances in Morphogenesis, Brachet J, King TJV eds. Academic Press, New York. 1968; 7: 205-249.
5. Dresden MH, Gross J. The collagenolytic enzyme of the regenerating limb of the newt Triturus viridescens. Dev Biol. 1970 May;22(1):129–157
6. Spotilla JR, Beumer RJ. The breeding habits of the ringed salamander, Ambystoma annulatum (Cope), in northwest Arkansas. Am Mid Natl. 1970; 84:77-89.
7. Toole BP, Gross J. The extracellular matrix of the regenerating newt limb: synthesis and removal of hyaluronate prior to differentiation. Dev Biol. 1971 May;25(1):57–77.
8. Scadding SR. Phylogenetic distribution of limb regeneration capacity in adult amphibia. J Exp Zool. 1972; 202:57-68.
9. Prichette WH, Dent JH. The role of size in the rate of limb regeneration in the adult newt. Growth. 1972; 36: 275-289.
10. Schauble MK. Seasonal variation of the newt forelimb regeneration under controlled environmental conditions. J Exp Zool. 1972; 181: 281-286.
11. Iten LE, Bryant. Forelimb regeneration from different levels of amputation in the newt, Notophthalamus viridescens: length, rate, stage. Wilhelm Roux Archiv. 1973; 173: 77-89.
12. Tank PW, Carlson BM, Connelly TG. A staging system for forelimb regeneration in the axolotl, Ambystoma mexicanum. J Morph. 1976; 150: 117-128.
13. Young HE. Limb Regeneration in the Adult Salamander, Ambystoma annulatum Cope 1889 (Amphibia: Ambystomatidae). University of Arkansas Library Press, copyright -1977;
14. Young HE. Epidermal ridge formation during limb regeneration in the adult salamander, Ambystoma annulatum. Proceedings of the Arkansas Academy of Science. 1977; 31:107-109;
15. Young HE, Bailey CF, Dalley BK. Environmental conditions prerequisite for complete limb regeneration in the postmetamorphic adult land-phase salamander, Ambystoma. Anatomical Record. 1983; 206:289-294;
16. Young HE, Bailey CF, Dalley BK. Gross morphological analysis of limb regeneration in postmetamorphic adult Ambystoma. Anatomical Record. 1983; 206:295-306;
17. Young HE. A Temporal Examination of Glycoconjugates During the Initiation Phase of Limb Regeneration in Adult Ambystoma. Texas Tech University Library Press, copyright 1983 and APPENDIX A Glycoconjugate Histochemistry Protocols, pp. 104-138.
18. Young HE, Dalley BK, Markwald RR. Identification of hyaluronate within peripheral nervous tissue matrices during limb regeneration. Edited by Coates, P.W., Markwald, R.R., Kenny, A.D., Alan R. Liss, Inc., New York. In: Developing and Regenerating Vertebrate Nervous Systems, Neurology and Neurobiology. 1983; 6:175-183;
19. Young HE, Bailey CF, Markwald RR, Dalley BK. Histological analysis of limb regeneration in postmetamorphic adult Ambystoma. Anatomical Record. 1985; 212:183-194.
20. Young HE, Speight MO. Allogeneic and autologous telomerase-positive stem cells as a potential treatment for systemic lupus erythematosus. Stem Cells Regen Med. 2020; 4(2):1-9.
21. Young HE, Young VE, Caplan AI. Comparison of fixatives for maximal retention of glycoconjugates for autoradiography, including use of sodium sulfate to release unincorporated radiolabeled [35S] sulfate. Journal of Histochemistry and Cytochemistry. 1989; 37:223-228.
22. Young HE, Carrino DA, Caplan AI. Histochemical analysis of newly synthesized and resident sulfated glycosaminoglycans during musculogenesis in the embryonic chick leg. Journal of Morphology. 1989; 201:85-103.
23. Young HE, Carrino DA, Caplan AI. Changes in synthesis of sulfated glycoconjugates during muscle development, maturation, and aging in embryonic to senescent CBF-1 mouse. Mechanisms of Ageing and Development. 1990; 53:179-193.
24. Young HE, Sippel J, Putnam LS, Lucas PA, Morrison DC. Enzyme-linked immuno-culture assay. Journal of Tissue Culture Methods. 1992; 14:31-36.
25. Young HE, Ceballos EM, Smith JC, Lucas PA, Morrison DC. Isolation of chick myosatellite and pluripotent stem cells. Journal of Tissue Culture Methods. 1992; 14:85-92.
26. Caplan AI. Mesenchymal stem cells. J Orthop Res. 1991; 9:641-650.
27. Young HE, Ceballos EM, Smith JC, Mancini ML, Wright RP, Ragan BL, Bushell I, Lucas PA. Pluripotent mesenchymal stem cells reside within avian connective tissue matrices. In Vitro Cellular & Developmental Biology. 1993; 29A:723-736.
28. Rogers JJ, Adkison LR, Black AC Jr, Lucas PA, Young HE. Differentiation factors induce expression of muscle, fat, cartilage, and bone in a clone of mouse pluripotent mesenchymal stem cells. The American Surgeon. 1995; 61(3):231-236.
29. Young HE, Mancini ML, Wright RP, Smith JC, Black AC Jr, Reagan CR, Lucas PA. Pluripotent mesenchymal stem cells reside within the connective tissues of many organs. Developmental Dynamics. 1995; 202:137-144.
30. Young HE, Speight MO. Characterization of endogenous telomerase-positive stem cells for regenerative medicine, a review. Stem Cell Regen Med 2020; 4(2):1-14.
31. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999 Apr 2;284(5411):143–147.
32. Pittenger MF, Discher DE, Péault BM, Phinney DG, Hare JM, Caplan AI. Mesenchymal stem cell perspective: cell biology to clinical progress. NPJ Regen Med. 2019 Dec 2; 4:22.
33. Gage FH. Mammalian neural stem cells. Science. 2000 Feb 25; 287(5457): 1433–1438.
34. Palmer TD, Takahashi J, Gage FH. The adult rat hippocampus contains primordial neural stem cells. Mol Cell Neurosci. 1997; 8(6): 389–404.
35. Temple S. The development of neural stem cells. Nature. 2001; 414: 112–117.
36. Götz M, Huttner WB. The cell biology of neurogenesis. Nat Rev Mol Cell Biol. 2005; 6(10): 777–788.
37. Ming G-L, Song H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron. 2011;70(4):687–702
38. Kriegstein A, Alvarez-Buylla A. The glial nature of embryonic and adult neural stem cells. Annu Rev Neurosci. 2009; 32: 149–184.
39. Urbán N, Guillemot F. Neural stem cells: generating and regenerating the brain. Neuron. 2014; 83(1): 18–36.
40. Baum CM, Weissman IL, Tsukamoto AS, Buckle AM, Peault B. Isolation of a candidate human hematopoietic stem-cell population. Proc Natl Acad Sci U S A. 1992 Apr 1; 89(7): 2804–2808.
41. Weissman IL, Shizuru JA. The origins of the identification and isolation of hematopoietic stem cells, and their capability to induce donor-specific transplantation tolerance and treat autoimmune diseases. Blood. 2008 Nov 1; 112(9): 3543–3553.
42. Seita J, Weissman IL. Hematopoietic stem cell: self-renewal versus differentiation. Wiley Interdiscip Rev Syst Biol Med. 2010; 2(6): 640–653.
43. Kent DG, Copley MR, Benz C, Wöhrer S, Dykstra BJ, Ma E, et al. Prospective isolation and molecular characterization of hematopoietic stem cells with durable self renewal potential.
Blood. 2009 Jun 18; 113(25): 6342–6350.
44. Kiel MJ, Yilmaz OH, Iwashita T, Yilmaz OH, Terhorst C, Morrison SJ. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell. 2005 Jul 1; 121(7): 1109–1121.
45. Laurenti E, Göttgens B. From haematopoietic stem cells to complex differentiation landscapes. Nature. 2018 Jan 18; 553(7689):418–426.
46. Wilkinson AC, Yamazaki S, Nakauchi H. Hematopoietic stem cells and their roles in tissue regeneration. Front Med (Lausanne). 2019; 6: 1–18.
47. Michalopoulos GK, Khan Z. Hepatic stem cells: in search of. Stem Cells. 2005; 23(6): 699 704.
48. Fausto N, Campbell JS, Riehle KJ. Liver regeneration. Hepatology. 2006; 43(2 Suppl 1): S45 53.
49. Turner R, Lozoya O, Wang Y, et al. Human hepatic stem cell and maturational liver lineage biology. Hepatology. 2011; 53(3): 1035 1045.
50. Petersen BE, Grossbard B, Goff JP, et al. The origin and liver repopulating capacity of murine oval cells. Proc Natl Acad Sci U S A. 2003; 100(Suppl 1): 11881 11888.
51. Fausto N, Campbell JS. The role of hepatocytes and oval cells in liver regeneration and repopulation. Mech Dev. 2003; 120(1): 117 130.
52. Tsamandas AC, Syrokosta I, Zolota V, et al. Oval cells in the liver: a putative stem cell compartment in chronic hepatitis and cirrhosis. Ann Gastroenterol. 2006; 19(3): 230 236.
53. Smukler SR, Arntfield ME, Razavi R, et al. The adult mouse and human pancreas contain rare multipotent stem cells that express insulin. Cell Stem Cell. 2011; 8(3): 281 293.
54. Seaberg RM, Smukler SR, Kieffer TJ, et al. Clonal identification of multipotent precursors from adult mouse pancreas that generate neural and pancreatic lineages. Stem Cells. 2004; 22(7): 1070 1083.
55. Xu X, D’Hoker J, Stangé G, et al. Beta cells can be generated from endogenous progenitors in injured adult mouse pancreas. Cell. 2008; 132(2): 197 207.
56. Inada A, Nienaber C, Katsuta H, et al. Carbonic anhydrase II–positive pancreatic cells are progenitors for both endocrine and exocrine pancreas after birth. Proc Natl Acad Sci U S A. 2008; 105(50): 19915 19919.
57. Dor Y, Brown J, Martinez OI, Melton DA. Adult pancreatic beta cells are formed by self duplication rather than stem cell differentiation. Nature. 2004; 429(6987): 41 46.
58. Hong KU, Reynolds SD, Watkins S, Fuchs E, Stripp BR. Basal cells are a stem cell population in the normal adult mouse trachea. Proc Natl Acad Sci U S A. 2004; 101(7): 2646 2651.
59. Kim CF, Jackson EL, Woolfenden AE, Lawrence S, Babar I, Vogel S, et al. Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell. 2005; 121(6): 823 835.
60. Giangreco A, Reynolds SD, Stripp BR. Terminal bronchioles harbor a unique airway stem cell population that localizes to the bronchioalveolar duct junction. Am J Pathol. 2002; 161(1): 173 182.
61. Engelhardt JF. Stem cells in the lung. In: Stripp BR, Reynolds SD, editors. Stem Cells in the Lung. New York: Marcel Dekker; 2005. p. 1 25.
62. Kajstura J, Rota M, Hall SR, Hosoda T, D’Amario D, Sanada F, et al. Identification of novel resident pulmonary stem cells: form and function of lung stem cells. Stem Cells. 2005; 23(8): 1073 1085.
63. Kotton DN, Fine A. Lung stem cells: new paradigms. Thorax. 2005; 60(3): 189 190.
64. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science. 1998; 6: 282(5391):1145-7.
65. Young HE, Black Jr AC. Adult Stem Cells. Anatomical Record Part A. 2004; 267A:75-102.
66. Nugent CI, Lundblad V. The telomerase reverse transcriptase: structure and function. Microbiol Mol Biol Rev. 2002;66(3):407–425.
67. Schmidt JC, Cech TR. Telomerase: mechanism of telomere synthesis. Annu Rev Biochem. 2012; 81:243–264.
68. Shammas MA. Telomeres, lifestyle, cancer, and aging. Curr Opin Clin Nutr Metab Care. 2011;14(1):28–34.
69. Shay JW, Wright WE. Telomeres and telomerase: three decades of progress. Nat Rev Genet. 2019;20(5):299–309
70. Smith AG, Heath JK, Donaldson DD, Wong GG, Moreau J, Stahl M, Rogers D. Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature. 1988; 336(6200):688–690.
71. Smith AG, Heath JK, Donaldson DD, Wong GG, Moreau J, Stahl M, Rogers D. Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature. 1988; 336(6200):688–690.
72. Niwa H, Burdon T, Chambers I, Smith A. Self-renewal of pluripotent embryonic stem cells is mediated via activation of STAT3. Genes Dev. 1998; 12(13):2048–2060.
73. Ohtsuka S, Dalton S. Regulation of embryonic stem cell self-renewal and pluripotency by the LIF signaling pathway. J Biochem. 2008; 144(6):713–718.
74. Onishi K, Zandstra PW. LIF signaling in stem cells and development. Development. 2015; 142(13):2230–2236.
75. Onishi K, Zandstra PW. LIF signaling in stem cells and development. Development. 2015; 142(13):2230–2236.
76. Humphrey RK, Beattie GM, Lopez AD, Bucay N, King CC, Firpo MT, et al. Maintenance of pluripotency in human embryonic stem cells is not dependent on LIF/STAT3 signaling. Stem Cells. 2004; 22(5):770–778.
77. Tang C, Lee AS, Volkmer JP, Sahoo D, Nag D, Mosley AR, et al. An antibody against SSEA 5 glycan on human pluripotent stem cells enables removal of teratoma forming cells. Nat Biotechnol. 2011; 29(9): 829 834.
78. Laflamme MA, Chen KY, Naumova AV, Muskheli V, Fugate JA, Dupras SK, et al. Cardiomyocytes derived from human embryonic stem cells in pro survival factors enhance function of infarcted rat hearts. Nat Biotechnol. 2007; 25(9): 1015 1024.
79. Kehat I, Kenyagin Karsenti D, Snir M, Segev H, Amit M, Gepstein A, et al. Human embryonic stem cells can differentiate into cardiomyocytes, exhibit functional properties of heart cells, and integrate into the rat heart. J Clin Invest. 2001; 108(3): 407 414.
80. Nussbaum J, Minami E, Laflamme MA, Virag JA, Ware CB, Masino A, et al. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. FASEB J. 2007; 21(7): 1345 1357.
81. Lee HJ, Lim IJ, Park SW, Kim YH, Nam HY, Jeon YJ, et al. Human embryonic stem cell derived neural precursors prevent teratoma formation by differentiation and withdrawal of pluripotent cells. Stem Cells Dev. 2012; 21(4): 594 602.
82. Lee AS, Tang C, Rao MS, Weissman IL, Wu JC. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat Med. 2013; 19(8): 998 1004.
83. Hentze H, Soong PL, Wang ST, Phillips BW, Putti TC, Dunn NR. Teratoma formation by human embryonic stem cells: evaluation of essential parameters for future safety studies. Stem Cell Res. 2009; 2(3): 198 210.
84. Moore KL, Persaud TVN, Torchia MG. The Developing Human: Clinically Oriented Embryology. 11th ed. Philadelphia: Elsevier; 2020. p. 25 56.
85. Schoenwolf GC, Bleyl SB, Brauer PR, Francis-West PH. Larsen’s Human Embryology. 6th ed. Philadelphia: Elsevier; 2021. p. 35 70.
86. Moore KL, Dalley AF, Agur AMR. Clinically Oriented Anatomy. 8th ed. Philadelphia: Wolters Kluwer; 2018.
87. Lee AS, Xu D, Plews JR, Nguyen PK, Nag D, Lyons JK, et al. Effects of cell number on teratoma formation by human embryonic stem cells. Cell Transplant. 2009; 18(12): 1249 1257.
88. Gertow K, Hirst CE, Stanley EG, Elefanty AG. Standardization of the teratoma assay for analysis of pluripotency of human ES cells and iPS cells. PLoS One. 2012; 7(9): e45532.
89. Hentze H, Soong PL, Wang ST, Phillips BW, Putti TC, Dunn NR. Teratoma formation by human embryonic stem cells: evaluation of essential parameters for future safety studies. Stem Cell Res. 2009; 2(3): 198 210.
90. Mueller Klieser W, Tonn T, Heidenreich O, et al. Teratomas derived from embryonic stem cells as models for tumor biology. In: Schatten H, editor. Teratomas Derived from Embryonic Stem Cells as Models for Tumorigenesis. Rijeka: IntechOpen; 2011. p. 1 23.
91. Reubinoff BE, Pera MF, Fong CY, Trounson A, Bongso A. Embryonic stem cell lines from human blastocysts: somatic differentiation in vitro and teratoma formation in vivo. Nat Biotechnol. 2000; 18(4): 399 404
92. Lucas PA, Calcutt AF, Southerland SS, Wilson JA, Harvey RL, Warejcka D, Young HE. A population of cells resident within embryonic and newborn rat skeletal muscle is capable of differentiating into multiple mesodermal phenotypes. Wound Repair and Regeneration. 1995; 3:449-460;
93. Warejcka DJ, Harvey R, Taylor BJ, Young HE, Lucas PA. A population of cells isolated from rat heart capable of differentiating into several mesodermal phenotypes. J Surg Res. 1996; 62:233-242;
94. Lucas PA, Warejcka DJ, Zhang L-M, Newman WH, Young HE. Effect of rat mesenchymal stem cells on the development of abdominal adhesions after surgery. J Surg Res. 1996; 62:229-232;
95. Lucas PA, Warejcka DJ, Young HE, Lee BY. Formation of abdominal adhesions is inhibited by antibodies to transforming growth factor-beta1. J Surg Res. 1996; 65:135-138;
96. Dixon K, Murphy RW, Southerland SS, Young HE, Dalton ML, Lucas PA. Recombinant human bone morphogenetic proteins-2 and 4 (rhBMP-2 and rhBMP-4) induce several mesenchymal phenotypes in culture. Wound Repair and Regeneration. 1996; 4:374-380;
97. Young HE, Wright RP, Mancini ML, Lucas PA, Reagan CR, Black AC Jr. Bioactive factors affect proliferation and phenotypic expression in pluripotent and progenitor mesenchymal stem cells. Wound Repair and Regeneration. 1998; 6(1):65-75;
98. Young HE, Rogers JJ, Adkison LR, Lucas PA, Black AC Jr. Muscle morphogenetic protein induces myogenic gene expression in Swiss-3T3 cells. Wound Rep Reg. 1998; 6(6):543- 554;
99. Young HE, Steele T, Bray RA, Detmer K, Blake LW, Lucas PA, Black AC Jr. Human progenitor and pluripotent cells display cell surface cluster differentiation markers CD10, CD13, CD56, CD90 and MHC Class-I. Proc Soc Exp Biol Med. 1999; 221:63-71;
100. Young HE. Pluripotent stem cells. Edited by M.A. Brown and S. Neufield, Cambridge Healthtech Institute Press, Newton Upper Falls, MA. In: Second Annual Symposium on Tissue Engineering / Regenerative Healing / Stem Cell Biology. 1999; 469-530;
101. Young HE. Stem cells and tissue engineering. In: Gene Therapy in Orthopaedic and Sports Medicine, J. Huard and F.H. Fu, eds., Springer-Verlag New York, Inc., Chap. 9, pg. 143-173, 2000;
102. Young HE, Duplaa C, Young TM, Floyd JA, Reeves ML, Davis KH, Mancini GJ, Eaton ME, Hill JD, Thomas K, Austin T, Edwards C, Cuzzourt J, Parikh A, Groom J, Hudson J, Black AC Jr. Clonogenic analysis reveals reserve stem cells in postnatal mammals. I. Pluripotent mesenchymal stem cells. Anat. Rec. 263:350-360, 2001.
103. Young HE, Steele T, Bray RA, Hudson J, Floyd JA, Hawkins K, Thomas K, Austin T, Edwards C, Cuzzourt J, Duenzl M, Lucas PA, Black AC Jr. Human reserve pluripotent mesenchymal stem cells are present in the connective tissues of skeletal muscle and dermis derived from fetal, adult, and geriatric donors. Anat. Rec. 264:51-62, 2001.
104. Romero-Ramos M, Vourc’h P, Young HE, Lucas PA, Wu Y, Chivatakarn O, Zaman R, Dunkelman N, El-Kalay MA, Chesselet M-F. Neuronal differentiation of stem cells isolated from adult muscle. J Neurosci Res 69:894-907, 2002.
105. Young HE. Existence of reserve quiescent stem cells in adults, from amphibians to humans. Curr Top Microbiol Immunol. 280:71-109, 2004.
106. Young HE, Duplaa C, Romero-Ramos M, Chesselet M-F, Vourc’h P, Yost MJ, Ericson K, Terracio L, Asahara T, Masuda H, Tamura-Ninomiya S, Detmer K, Bray RA, Steele TA, Hixson D, El-Kalay M, Tobin BW, Russ RD, Horst MN, Floyd JA, Henson NL, Hawkins KC, Groom J, Parikh A, Blake L, Bland LJ, Thompson AJ, Kirincich A, Moreau C, Hudson J, Bowyer III FP, Lin TJ, Black Jr AC. Adult reserve stem cells and their potential for tissue engineering. Cell Biochem Biophys, 40(1):1-80, 2004.
107. Young HE, Duplaa C, Yost MJ, Henson NL, Floyd JA, Detmer K, Thompson AJ, Powell SW, Gamblin TC, Kizziah K, Holland BH, Boev A, Van de Water JM, Godbee DC, S. Jackson, M. Rimando, Edwards CR, Wu E, Cawley C, Edwards PD, Macgregor A, Bozof R, Thompson TM, Petro Jr GJ, Shelton HM, McCampbell BL, Mills JC, Flynt FL, Steele TA, Kearney M, Kirincich-Greathead A, Hardy W, Young PR, Amin AV, Williams RS, Horton MM, McGuinn S, Hawkins KC, Ericson K, Terracio L, Moreau C, Hixson D, Tobin BW, Hudson J, Bowyer III FP, Black Jr AC. Clonogenic analysis reveals reserve stem cells in postnatal mammals. II. Pluripotent epiblastic-like stem cells. Anat. Rec. 277A:178-203, 2004.
108. Vourc'h P, Romero-Ramos M, Chivatakarn O, Young HE, Lucas PA, El-Kalay M, Chesselet M-F. Isolation and characterization of cells with neurogenic potential from adult skeletal muscle. Biochemical and Biophysical Research Communications 317:893-901, 2004.
109. Seruya M, Shah A, Pedrotty D, du Laney T, Melgiri R, McKee JA, Young HE, Niklason LE. Clonal Population of adult stem cells: life span and differentiation potential. Cell Transplant 13:93-101, 2004
110. Young HE, Black AC Jr. Differentiation potential of adult stem cells. In: Contemporary Endocrinology: Stem Cells in Endocrinology, L.B. Lester, ed., The Humana Press Inc., Totowa, NJ. Chap. 4, p. 67-92, 2005b.
111. Vourc’h P, Lacar B, Mignon L, Lucas PA, Young HE, Chesselet MF. Effect of neurturin on multipotent cells isolated from the adult skeletal muscle. Biochem Biophys Res Commun 332:215-223, 2005.
112. Henson NL, Heaton ML, Holland BH, Hawkins KC, Rawlings B, Eanes E, Bozof R, Powell S, Grau R, Fortney J, Peebles B, Kumar D, Yoon JI, Godby K, Collins JA, Sood R, Bowyer 3rd FP, Black Jr AC, Young HE. Karyotypic analysis of adult pluripotent stem cells. Histology and Histopathology, 20: 769-784, 2005.
113. Mignon L, Vourc'h P, Romero-Ramos M, Osztermann P, Young HE, Lucas PA, Chesselet MF. Transplantation of multipotent cells extracted from adult skeletal muscles into the adult subventricular zone of adult rats. J Comp Neurol 491:96-108, 2005.
114. Young HE, Duplaa C, Katz R, Thompson T, Hawkins KC, Boev AN, Henson NL, Heaton M, Sood R, Ashley D, Stout C, Morgan JH, Uchakin PN, Rimando M, Long GF, Thomas C, Yoon JI, Park JE, Hunt DJ, Walsh NM, Davis JC, Lightner JE, Hutchings AM, Murphy ML, Boswell E, McAbee JA, Gray BM, Piskurich J, Blake L, Collins JA, Moreau C, Hixson D, Bowyer FP, Black AC Jr. Adult-derived stem cells and their potential for tissue repair and molecular medicine. J Cell Molec Med 9:753-769, 2005.
115. Young HE, Black AC Jr. Adult-derived stem cells. Minerva Biotechnologica Cancer Gene Mechanisms and Gene Therapy Reviews 17:55-63, 2005.
116. Stout CL, Ashley DW, Morgan III JH, Long GF; Collins JA, Limnios JI, Lochner F, McCommon G, Hixson D, Black Jr AC, Young HE. Primitive stem cells reside in adult swine skeletal muscle and are mobilized into the peripheral blood following trauma. American Surgeon 73 (11):1106-1110, 2007.
117. Stout CL, McKenzie J, Long G, Henson N, Hawkins KC, Ashley DW, Collins J, Hixson D, Black Jr AC, Young HE. Discovery of pluripotent and totipotent stem cells in the heart of the adult rat. Amer Surg 73: S63, 2007.
118. Vourc’h P, Mignon L, Lucas PA, Young HE, Chesselet MF. Cells isolated from adult skeletal muscle express markers of differentiated neurons after transplantation into the adult hippocampus. (In press).
119. Young HE and Black Jr AC. Naturally occurring adult pluripotent stem cells. In: Stem Cells: From Biology to Therapy, Advances in Molecular Biology and Medicine. 1st Ed, R.A. Meyers, Ed, WILEY-BLACKWELL-VCH Verlag GmbH & Co. KGaA. Chap 3, pp. 63-93, 2013.
120. McCommon GW, Lochner F, Black Jr AC, Young HE. Primitive adult-derived stem cells are present in the blood of adult equines and can be increased in number with moderate exercise or ingestion of a cyanobacter, Aphanizomenon flos-aquae. Autocoids 2: 103, 2013,
121. Young HE, Henson NL, Black GF, Hawkins KC, Coleman JA, Black Jr AC. Location and characterization of totipotent stem cells and pluripotent stem cells in the skeletal muscle of the adult rat. J Stem Cell Res 1(1) 002: 1-17, 2017.
122. Young HE, Lochner F, Lochner D, Lochner D, McCommon G, Black AC Jr. Primitive Stem Cells in Adult Feline, Canine, Ovine, Caprine, Bovine, and Equine Peripheral Blood. J Stem Cell Res. 1(1) 004: 1-6, 2017.
123. Young HE, Lochner F, Lochner D, Lochner D, Black GF, Coleman JA, Young VE, McCommon G, Black Jr AC. Primitive stem cells in adult human peripheral blood. J Stem Cell Res. 1(2) 001:1-8, 2017.
124. Young HE, Henson NL, Black GF, Hawkins KC, Coleman JA, Black Jr AC. Stage-Specific Embryonic Antigen-4-Positive Cells and Carcinoembryonic Antigen Cell Adhesion Molecule-1-Positive Cells are Located in the Bone Marrow of the Adult Rat. J Stem Cell Res. 1(2) 001: 1-3, 2017.
125. Young HE, Limnios JI, Lochner F, McCommon G, Black GF, Coleman JA, Hawkins KC, Black Jr AC. Healing cells in the dermis and adipose tissue of the adult pig. J Stem Cell Res 2017; 1(2) 004:1-5.
126. Young HE, Black GF, Coleman JA, Hawkins KC, Williams S, Black Jr AC. Healing cells in the kidney of the adult rat. J Stem Cell Res 2017; 1(3) 001:1-4.
127. Young HE, Limnios JI, Lochner F, McCommon G, Black GF, Coleman JA, Hawkins KC, Black Jr AC. Healing cells in the dermis and adipose tissue of the adult pig. J Stem Cell Res 2017; 1(2) 004:1-5.
128. Young HE, Lochner F. Telomerase positive totipotent stem cells in the adult brain. I. cerebral cortex. Regen Med Biol Res 2021; 2(1):1-16.
129. Young HE, Lochner F. Endogenous adult telomerase positive stem cells increase in equine peripheral blood following exercise. J Reg Med Biol Res. 2024; 5(2):1-8.
130. Young HE. Carcinoembryonic antigen-cell adhesion molecule-1 and stage-specific embryonic antigen-4 are present in the reproductive organs of adult mammals. GSC Advanced Research and Reviews. 2025; 23(03): 149-157.
131. Young HE. Totipotent stem cells and pluripotent stem cells are present in the reproductive organs of an adult mammal. GSC Advanced Research and Reviews. 2025; 23(03): 158-180.
132. Yamanaka S. A fresh look at iPS cells. Cell. 2009;137(1):13 17.
133. Takahashi K, Narita M, Yokura M, Ichisaka T, Yamanaka S. Human induced pluripotent stem cells on autologous feeders. PLoS One. 2009; 4(12): e8067.
134. Ohnuki M, Takahashi K, Yamanaka S. Generation and characterization of human induced pluripotent stem cells. Curr Protoc Stem Cell Biol. 2009; Chapter 4: Unit 4A.2.
135. Armanios M, Blackburn EH. The telomere syndromes. Nat Rev Genet. 2012; 13(10): 693 704.
136. Blackburn EH, Epel ES, Lin J. Human telomere biology: a contributory and interactive factor in aging, disease risks, and protection. Science. 2015; 350(6265): 1193 1198.
137. Lansdorp PM. Telomeres and telomerase: a simple end matter. Nat Med. 2005;11(10):1049 1054.
138. Schmidt JC, Cech TR. Human telomerase: biogenesis, trafficking, recruitment, and activation. Genes Dev. 2015; 29(11): 1095 1105.
139. Kordinas V, Tsirpanlis G, Imprialos K, Giannopoulou E, Doumas M. Telomere length, telomerase activity and atherosclerosis. Curr Med Chem. 2016; 23(12): 1280 1290.
140. Batista LFZ, Artandi SE. Telomere and telomerase dynamics in induced pluripotent stem cells. World J Stem Cells. 2013; 5(3): 116 123.
141. Ahmed MS, Ikram S, Bibi N, Mir A. Telomere biology in induced pluripotent stem cells: implications for aging and regenerative medicine. World J Stem Cells. 2016; 8(6): 182 194.
142. Jenkins YM, de Lange T. Telomerase and induced pluripotency. Cell Res. 2012; 22(4): 614 617.
143. Wang J, Xie LY, Allan S, Beach D, Hannon GJ. Myc activates telomerase. Genes Dev. 1998; 12(12): 1769 1774.
144. López de Si.lanes I, Graña O, De Bonis ML, Dominguez O, Pisano DG, Blasco MA. Identification of TERRA locus unveils a telomerase regulating role for telomeric RNAs in iPSC. Cell. 2014; 158(6): 1232 1245.
145. Tomov ML, Yamada S, Pace J, et al. iPSC culture: what every researcher needs to know. Tempo Biosci Technical Bulletin. 2025; 1: 1 12.
146. JangoCell Scientific Team. Detecting and preventing spontaneous differentiation in iPSC and MSC cultures. JangoCell Application Note. 2025; 1: 1 10.
147. Stoilova T, Hristova A, Kolev M, et al. Improving the differentiation potential of pluripotent stem cells by controlling spontaneous differentiation. Sci Rep. 2022; 12(1): 14321.
148. Chambers SM, Fasano CA, Papapetrou EP, Tomishima M, Sadelain M, Studer L. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Proc Natl Acad Sci U S A. 2009; 106(33): 12741 12746.
149. Gertow K, Hirst CE, Stanley EG, Elefanty AG. Teratoma formation: a tool for monitoring pluripotency in stem cell research. Curr Protoc Stem Cell Biol. 2015; 32: 4A.8.1 4A.8.23.
150. Ben David U, Benvenisty N. The tumorigenicity of human embryonic and induced pluripotent stem cells. Nat Rev Cancer. 2011; 11(4): 268 277.
151. Young HE, Black AC. Pluripotent Stem Cells, Endogenous versus Reprogrammed, a Review. MOJ Orthop Rheumatol 1(4): 00019, 2014.
152. Young HE, Black GF, Coleman JA, Hawkins KC, Black Jr AC. Pulmonary diseases and adult healing cells: from bench top to bedside. J Stem Cell Res 2017; 1(2) 003:1-9.
153. Young HE, Hyer L, Black AC Jr, Robinson Jr JS. Treating Parkinson disease with adult stem cells. J Neurological Disorders, 2:107-109, 2013b.
154. Young HE, Hyer L, Black AC Jr, Robinson Jr JS. Adult stem cells: from bench-top to bedside. In: Tissue Regeneration: Where Nanostructure Meets Biology, 3DBiotech, North Brunswick, NJ Chap 1, pp 1-60, 2013a.
155. Young HE, Limnios IJ, Lochner F, McCommon G, Black GF, Coleman JA, Hawkins KC, Black Jr AC. Cardiovascular disease and adult healing cells: From bench top to bedside. J Stem Cell Res 2017; 1(3) 002:1-8.
156. Black Jr AC, Williams S, Young HE. From Bench Top to Bedside: Formation of Pulmonary Alveolar Epithelial Cells by Maintenance Cells and Healing Cells. J Stem Cell Res. 2017; 1(2) 002: 1-16.
157. Young HE, Speight MO. Local Anesthetics Can Affect the Efficacy of Telomerase-Positive Stem Cells. J Regen Med. Biol Res 2020; 1(1):1-9.
158. Young HE, Speight MO. Osteoarthritis Treated with Telomerase-Positive Adult Stem Cells in Animals and Humans. Stem Cells Regen Med. 2020; 4(2):1-11. JSCR-20-049.
159. Young HE, Speight MO. Informed consent guidelines for optimizing the use of telomerase-positive stem cells for regenerative medicine. J Regen Med Biol Res 2020; 1(1):1-20.
160. Young HE, Speight MO. Telomerase-positive stem cells as a potential treatment for idiopathic pulmonary fibrosis. Stem Cells Regen Med. 2020; 4(2):1-11.
161. Young HE, Speight MO. Criteria to Distinguish TSCs from Exosomes as Major Players in Regenerative Medicine. J Regen Med & Biol Res. 2020; 1(1):1-5. JRMBR-1(1)-005.
162. Young HE, Speight MO. Potential treatment of chronic obstructive pulmonary disease with allogeneic and autologous telomerase-positive stem cells. Stem Cells Regen Med. 2020; 4(3):1-11. JSCR-20-056.
163. Young HE, Speight MO. Allogeneic telomerase-positive stem cells as a treatment for celiac disease. Stem Cells Regen Med. 2020; 4(2):1-7 JSCR-20-050.
164. Young HE, Speight MO. Cardiovascular disease treated with telomerase-positive stem cells. Stem Cells Regen Med. 2020; 4(2):1-8 JSCR-20-051.
165. Young HE, Speight MO. Age-related macular degeneration treated with autologous telomerase-positive totipotent stem cells. Stem Cells Regen Med. 2020; 4(3):1-9. JSCR-20-055.
166. Young HE, Speight MO. Donor selection is a critical component using naïve endogenous adult stem cells for the treatment of chronic diseases and traumatic injuries. J Regen Med & Biol Res. 2020; 1(2):1-28. JRMBR-1(2)-009.
167. Young HE, Speight MO. Alzheimer’s disease treated with autologous and allogeneic telomerase-positive stem cells. Stem Cells & Regen Med. 2021; 5(1):1-17.
168. Young HE, Speight MO. Blunt force trauma-induced total bilateral vision impairment of 13 years duration treated with autologous telomerase positive stem cells. Stem Cells Regen Med. 2021; 5(1):1-22.
169. Young HE, Speight MO. Traumatic spinal cord injury treated with autologous telomerase-positive stem cells. Stem Cells Regen Med. 2021; 5(1):1-13.
170. Young HE, Speight MO. Chronic inflammatory demyelinating polyneuropathy treated with autologous telomerase-positive stem cells. Stem Cells Regen Med. 2021; 5(2):1-16.
171. Young HE, Speight MO. Treating Parkinson Disease with Autologous Telomerase-Positive Stem Cells, Update 2021. Stem Cells & Regenerative Medicine. 2021; 5(1): 1-13.
172. Young HE. Healing Cells: Use What the Almighty Created to Heal Thyself. American Journal of Medical and Clinical Research & Reviews. 2023; 2(7):1-19.
173. Young HE. A high throughput screening assay to quantify, visualize, and standardize biological activities: Enzyme-Linked Immuno-Culture Assay (ELICA). GSC Advanced Research and Reviews. 2025; 24(02): 091-114.
174. Young HE, Speight MO. Characterization of endogenous telomerase-positive stem cells for regenerative medicine, a review. Stem Cell Regen Med 2020; 4(2):1-14.
175. Young HE. Endogenous Adult Telomerase Positive Stem Cells (aTPSCs) and Combinatorial Nutraceutical Supplement Pill (CNSP), 50+ Years in the Making, 50+ Years of Discovery; In preparation.
176. Caplan AI. Mesenchymal stem cells: Time to change the name! Stem Cells Transl Med. 2017; 6(6): 1445–1451.
177. Caplan AI, Correa D. The MSC: an injury drugstore. Cell Stem Cell. 2011; 9(1): 11–15
178. Caplan AI, Dennis JE. Mesenchymal stem cells as trophic mediators. J Cell Biochem. 2006 Aug 1; 98(5): 1076–1084.
179. Dominici M, Le Blanc K, Mueller I, Slaper Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. Cytotherapy. 2006;8(4):315 317.
180. Lv F J, Tuan RS, Cheung KMC, Leung VYL. Concise review: the surface markers and identity of human mesenchymal stem cells. Stem Cells. 2014;32(6):1408 1419.
181. Samsonraj RM, Raghunath M, Nurcombe V, Hui JH, van Wijnen AJ, Cool SM. Concise review: multifaceted characterization of human mesenchymal stem cells for use in regenerative medicine. Stem Cells Transl Med. 2017;6(12):2173 2185.
182. Kuci S, Kuci Z, Schäfer R, Bruderek K, Schardt A, Blaeschke F, et al. Clonal analysis of multipotent stromal cells reveals limited hierarchies. Stem Cells. 2018;36(12):1909 1924.
183. Phinney DG, Pittenger MF. Concise review: MSC derived exosomes for cell free therapy. Stem Cells. 2017;35(4):851 858.
184. Young HE, Speight MO, Black AC Jr. Functional Cells, Maintenance Cells, and Healing Cells. J Stem Cell Res. 1(1): 003: 1-4, 2017.
185. Zvereva MI, Shcherbakova DM, Donsta OA. Telomerase: structure, functions, and activity regulation. Biochemistry (Mosc.) 2010; 75: 1563-1583.
186. Young HE. Combinatorial Nutraceutical Supplement Pill (CNSP) Stimulates Naïve Adult Telomerase Positive Stem Cells In-Situ to Heal Cardiomyopathies. GSC Advanced Research and Reviews. 2024; 20(02), 047-056.
187. Young HE. Fresh Isolate Adult Telomerase Positive Stem Cells: An addition to Embryonic Stem Cells (ESCs), Induced Pluripotent Stem Cells (iPSCs), and/or Mesenchymal Stem Cells (MSCs) for Regenerative Medicine. GSC Advanced Research and Reviews. 2023; 16(1):066-081.
188. Young HE. Adult telomerase positive stem cells: isolation, plating, and propagation. GSC Advanced Research and Reviews. 2025; 25(02):407-440.
189. Young HE, Limnios JI, Lochner F, McCommon G, Cope LA, Black AC Jr. Pancreatic islet composites secrete insulin in response to a glucose challenge. J Stem Cell Res 1(1) 001: 1-12, 2017.
190. Hayflick L. The limited in vitro lifetime of human diploid cell strains. Exp Cell Res. 1965;37(3):614 636.
191. Kumar V, Abbas AK, Aster JC. Robbins Basic Pathology. 10th ed. Philadelphia: Elsevier; 2018.
192. Young HE. Adult telomerase positive stem cells: introduction and location. GSC Advanced Research and Reviews. 2025; 25(02): 296-331.
193. Young HE. Adult telomerase positive stem cells: isolation, plating, and propagation. GSC Advanced Research and Reviews. 2025; 25(02):407-440.
194. Young HE. Adult telomerase positive stem cells: differential cryopreservation, cell surface marker profiles, and cell sorting. MOJ Orthopedics & Regeneration. 2025; 17(5): 155-173.
195. Young HE. Adult telomerase positive stem cells: Cell-Specific Exosome-Conditioned Medium, Repetitive Single Cell Clonogenic Analysis, and Genomic Labeling. GSC Advanced Research and Reviews. 2025; 25(2): 480-504.
196. Young HE. Adult telomerase positive stem cells: Effects of Biological Agents on Single Cell Clones of Adult Cells. GSC Advanced Research and Reviews. 2025; 25(03): 028-047.
197. Young HE. Adult Telomerase Positive Stem Cells: Effects of Biological Agents on Genomically-Labeled Clones and Unlabeled Clones of Adult Cells. GSC Advanced Research and Reviews. 2025; 25(03): 127-145.
198. Young HE. Adult Telomerase Positive Stem Cells: Compare and Contrast Biobanking with Mesenchymal Stem Cells and Other Progenitor Cells. GSC Advanced Research and Reviews. 2025; 25(03): 218-226.
199. Young HE. Adult Telomerase Positive Stem Cells: Induced Proliferation of Precursor Cells by Platelet-Derived Growth Factor-BB. J Stem Cell Res. 2026; 7(1):1-15.
200. Young HE. Adult Telomerase Positive Stem Cells: Remain constant throughout lifespan of individual. J Stem Cell Res. 2026; 7(1):1-13.
201. Young HE. Combinatorial nutraceutical supplement pill (CNSP) stimulates naïve adult telomerase positive stem cells in-situ to reverse signs and symptoms in multiple health conditions. GSC Advan Res Rev. 2024; 20(02): 047-056.
No Files Found




Internationally Accepted
Share Your Publication :